

Diversity of psammophyte communities on sand dunes and sandy soils of the northern Sahara desert
⁎Corresponding author at: Department of Forest Management, Higher National School of Forests, 40000 Khenchela, Algeria chenchouni@gmail.com (Haroun Chenchouni),
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
-
Composition and diversity of plant communities of dune and sandy habitats in the Sahara was analyzed.
-
Plant functional traits (e.g. lifeforms, dispersal and Noy-Meir’s strategies) of Psammophyte communities were examined.
-
Psammophytic flora included 29 species, 27 genera, 18 families, predominated by Poaceae and Brassicaceae.
-
Plant diversity of Saharan sandy habitats was very low, dominanted by Stipagrostis pungens and S. acutiflora.
-
Flora mainly comprised perennials, chamaephytes, hemicryptophytes, anemochores and arido-active species.
Abstract
Background
Hot deserts contain several habitats where the diversity, structure and functional features of plant communities in certain habitats, such as sandy soils and dunes for instance, are little explored.
Objectives
This study aimed to characterize the flora of sandy soils of the northern Algerian Sahara in order to determine the different plant functional traits and conservation aspects of the psammophytic communities for a sustainable management of phytogenetic resources.
Methods
In seven sites representing different sandy habitats, spontaneous psammophytic communities were sampled using 70 floristic relevés ‘samples’ with an area of 100 m2 for each and 10 replicates per sampling site. In each sample, the number of individual of plant species was measured, then plant diversity and similarity was estimated per site. Several plant traits including lifeforms, morphological types, dispersal forms, Noy-Meir types and chorological categories were attributed for each plant species to characterize psammophyte communities.
Results
Overall, plant diversity of Saharan sandy habitats was very low. The sampled sandy soils and dunes included 29 plant species classified into 27 genera and 18 families, with good representation of Poaceae and Brassicaceae. The arido-active species were dominant with 62.1% of the flora, which were mainly represented by perennial plants belonging to chamaephytes, hemicryptophytes and phanerophytes. Plants of sandy soils and dunes adopted various dispersal types, mainly dominated by anemochorous species. The chorological analysis revealed the existence of two endemic species to the Sahara Desert whereas the Mediterranean-Saharan-Arabian species were mostly represented by the Saharo-Arabian element which adapts well to environmental conditions of the Sahara. The assessment of plant abundances showed that the community of psammophytes contained three rare plants, one quite rare species and two fairly common plants in the flora of Algeria. Accordingly, sandy habitats require adequate management measures in order to preserve the biodiversity in Saharan environments. The number of plant species, genera and families significantly increased with the increase in vegetation cover on sand dunes.
Conclusions
The results of this survey can represent a first floristic database providing good indicators on plant communities, habitat and environmental conditions of sandy soils and dunes of the Algerian Sahara.
Keywords
Algerian Sahara
Desert sand dune flora
Plant community
Plant diversity
Psammophyte
Sandy soils
Vegetation traits

1 Introduction
The Sahara Desert stretches across the northern third of the African continent from the Atlantic Ocean to the Red Sea (Le Houerou, 1990). Due to very low and irregular precipitation, high temperatures, large temperature range and long droughts, this ecoregion is characterized by severe environmental conditions that limit the survival of many living beings (Le Houerou, 1990; Ozenda, 2004). Still, several species survive and present extraordinary adaptations to these harsh environmental conditions. The distribution of spontaneous vegetation is mainly related to the geomorphological forms of this ecoregion including river “Wadi”, desert rocky plateaus “Hammada”, closed depressions “Daya”, desert pavements “Reg”, sandy soils and dunes, mountains “Djebel”, salty soils “Chott and Sabkha” and rocky soils (Benhouhou et al., 2001; Benhouhou et al., 2003; Ozenda, 2004; Chehma, 2005; Bouallala, 2013; Bradai et al., 2015a; Bouallala et al., 2020). Annual rainfall is very low but is sufficient to maintain plant life on almost all landscapes in the northern Sahara Desert. A landscape-specific vegetation is thus consisting of very uneven density and structure that depend on environmental characteristics of the habitat. The vegetation is denser in depressions such as Wadi beds, while scattered vegetation with low canopy cover occurs on desert plateaus, pavements and sand dunes (Bouallala et al., 2022; Ozenda, 2004).
Saharan spontaneous vegetation is quite diverse and has several uses, particularly as forage of grazing dromedaries (Longo-Hammouda et al., 2007; Bouallala et al., 2013; Bouallala et al., 2011; Chehma and Huguenin, 2017), as well as in the traditional pharmacopoeia for the treatment of various pathologies using a range of plant bioactive compounds (Codou-David, 2012; Hammiche and Maiza, 2006; Bensizerara et al., 2013; Bradai et al., 2020). Previous floristic surveys of habitats of the Sahara Desert, including sand soils, applied extensive plant morphological descriptors, such as Raunkiaer lifeforms and lifecycle/growth types, and chorological types to characterize plant communities (Quézel and Santa, 1962; 1963; Quézel, 1965; Ozenda, 2004; Bouallala, 2013; Médail and Quézel, 2018). Recent studies in some Saharan and sub-desertic regions applied more comprehensive characterizing approaches involving complementary plant functional traits i.e. dispersal types, Noy-Meir adaptations and Grime's CSR strategies (Bradai et al., 2015a; Neffar et al., 2018; Bouallala et al., 2020).
Despite the existence of some studies that aimed to the conservation of Saharan flora and vegetation in Algeria (Barry et al., 1970; Abdelkrim, 2006; Médail and Quézel, 2018), plant communities of specific habitats such as sandy soils and sand dunes are still poorly studied. These types of hot arid habitats are considered very fragile ecosystems because of sand encroachment inducing the instability of the sand substrate and resulting in low habitat productivity (Bouallala et al., 2022). The characterization of plant communities using plant functional traits provides an accurate and reliable tool to understand vegetation-environment relationships in a changing world (Chenchouni, 2017; Kouba et al., 2021). The use of such kind of plant characterization approach is particularly needed in arid and semiarid lands where the vegetation is under environmental pressure due to climate change and severe drought events combined with increasing anthropogenic activities such as free grazing and/or unsustainable uses of rare and valuable plants for various purposes (Macheroum et al., 2021; Mehalaine and Chenchouni, 2021). In this context, this study aims at analyzing plant diversity of different sandy habitats in the northern Sahara Desert of Algeria in order to detect rare plant species and then suggest adequate conservation measures for plant community development and management plans for the conservation of hyper-arid ecosystems. The study aimed to provide a comprehensive analysis of plant community diversity using some biodiversity parameters such as species richness, Shannon diversity index and evenness and also qualitative and quantitative estimators of diversity similarity (beta diversity). We analyzed the taxonomic structures of plant assemblages using taxonomic ratios of the family or generic richness to species richness, and explored the relationships between the psammophytic vegetation cover and the richness of different taxa. This survey investigated plant community characteristics using some plant functional traits, namely Raunkiaer lifeforms, morphological types, dispersal forms, Noy-Meir types and chorological categories, and the appreciation of the abundance of psammophyte communities which remain very little studied.
2 Materials and methods
2.1 Study area
The study area is located in the northern part of the Algerian Sahara in the regions of Ghardaïa and Ouargla (Fig. 1). Plant communities in this area are well adapted to ecological conditions of the Saharan habitats: Hamada, Regs, sandy soils, salty soils, depressions, Wadi beds. The natural and wild-grown vegetation offers the main fodder resources for the dromedary farmed in extensive husbandry and free grazing system (Chehma et al., 2008a; b). The climate is hot hyperarid ‘desertic’ with a large inter- and intra-annual variability of precipitation (occurring exclusively as rainfall), high temperatures, large temperature range, very intense evapotranspiration, severe and long drought episodes, and extreme weather events such as heat waves and extreme wind events (Quézel, 1965; Ozenda, 2004; Bradai et al., 2015b). According to Köppen climate classification, the region had an arid climate of hot desert “BWh”. De Demartone aridity index varied between 1 and 2, indicating hyperarid climates with precipitation deficit ranged between 85 and 98% (Table S1). According to ombrothermic diagram of Bagnouls and Gaussen, the drought period extended over 12 months a year (Fig. 1). Rainfall was very low (<36–76 mm/year), with <12 days of effective rainy days and the potential evapotranspiration very significantly exceeded precipitations all year round. July was the hottest month with the average of maximum temperature exceeded 41 °C, whereas the minimum temperatures in January did not drop below 3.2–4.4 °C. Monthly sunshine frequency was higher than 70% (Table S2).
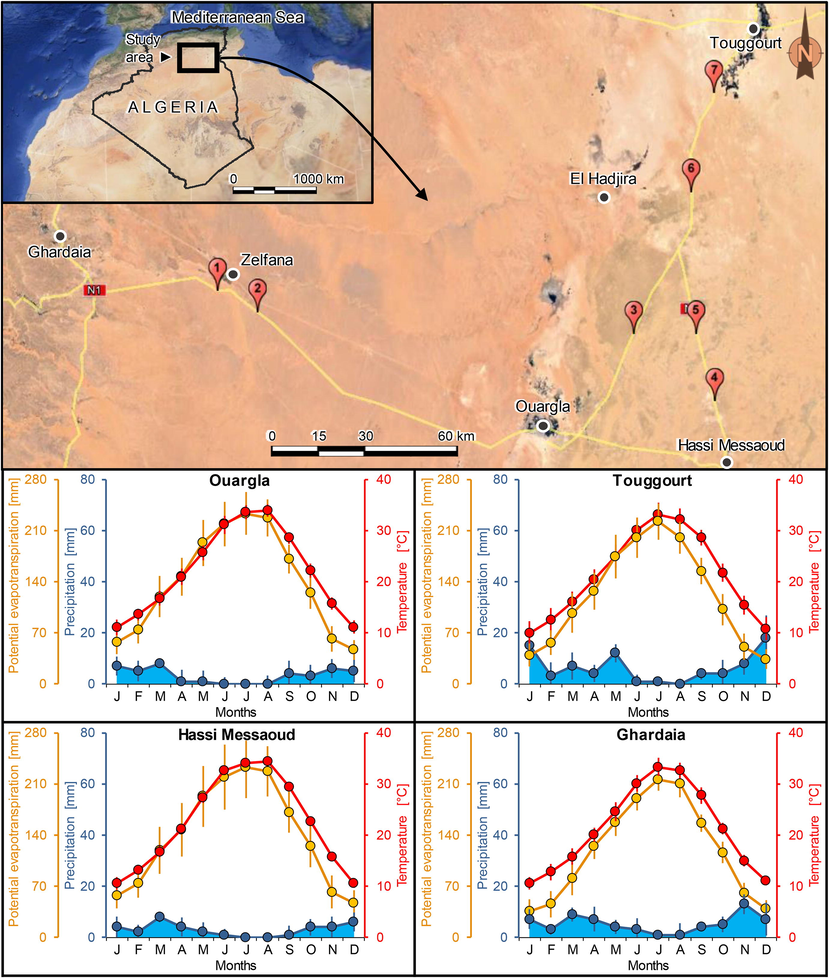
- Geographic map showing the location of sampling sites (numbered red pin icons) in sandy habitats of the Algerian Sahara Desert. Lower plots are ombrothermic diagrams of Bagnouls and Gaussen applied to meteorological data of Ouargla, Touggourt, Hassi Messouad, and Ghardaia.
2.2 Choice of sampling sites in sand habitats
The study area was visited several times prior choosing the sampling sites to study the diversity of spontaneous plant communities in sandy habitats. Seven sites (Fig. 1) were selected based on landscape geomorphology, shape of sand accumulation, and homogeneity of ecological conditions. Site 1 (altitude = 359.64 m above sea level) represents a micro-dune established on Wadi beds where Stipagrostis pungens dominates, site 2 (altitude = 354.62 m a.s.l.) is a sand-filled rocky-bottom Wadi bed with S. pungens as characteristic species of the vegetation, site 3 (altitude = 145 m a.s.l.) is a silted Reg with the dominance of Stipagrostis acutiflora, site 4 (altitude = 160 m) is a silted Reg dominated by S. pungens, site 5 (altitude = 132 m) is a silted Reg characterized by S. acutiflora, site 6 (altitude = 121 m a.s.l.) is a silted Reg dominated by S. acutiflora, and site 7 (altitude = 106 m a.s.l.) is a micro-dune where S. pungens is abundant.
2.3 Plant sampling and data collection
Assuming the homogeneity of environmental conditions within each site, ten sample plots were placed randomly in each of the selected sampling sites. Floristic surveys were conducted during the optimal period of vegetation development that coincides with favorable temperature and seasonal rainfall in the Sahara Desert (February-April 2015). The size of every plot of the ten samples was 100 m2 (10 m × 10 m), which is a sufficient sampling area given the scatterness of vegetation in hot desert environments, and sand habitats in particular. This sampling area is also adopted in most studies of Saharan flora and vegetation, which makes comparison with previous studies more reliable (Benhouhou et al., 2003, Bouallala, 2013). In each plot, the density of each species was determined. Plant species were identified using identification books of Algerian flora (Quézel and Santa, 1962; 1963; Ozenda, 2004).
2.4 Analysis of plant diversity
The floristic richness and diversity was assessed for each sampling site using several descriptors of alpha diversity (species richness, Shannon diversity index and evenness) and beta diversity (Jaccard index and Bray-Curtis distance). The data from the sample plots were treated as separate data entities when computing alpha diversity whereas they were lumped together for computing beta diversity indices using sums of species abundances from sample plots. In addition, the species richness recorded at the sampling sites was compared using species richness scale adopted in the Saharan regions (Daget and Poissonet, 1991). Accordingly, for each site, the status of the flora was determined following the number of species (S) as: very species-poor (S < 11), species-poor (S = 11–20), moderate (S = 21–30), fairy species-rich (S = 31–40), species-rich (S = 41–50) and very species-rich flora (S > 50).
Plant alpha diversity was measured using Shannon diversity index (H'),
Using the ratios of the generic or family richness to species richness (G/S or F/S), the phylogenetic diversity was analyzed through the taxonomic structures for each sampling site. Since the taxonomic structure of communities varies following habitat environmental conditions, the relationships between vegetation cover measured at each sampling plot and the number of families, genera and species were explored using Pearson correlation tests. We assumed that with the more favorable ecological conditions in sandy habitats, vegetation cover and accordingly the richness of hierarchical taxonomic categories (family, genus and species) would increase. Thus we predicted significant positive correlations between vegetation cover and the number of taxa at different taxonomic levels. The species-higher taxon relationships, i.e. relationships between species richness and generic or family richness, were investigated using power models linearized after natural logarithm (ln) transformations (Fan et al., 2017). We applied these models: ln(G) = a × ln(S) and ln(F) = a × ln(S) for the species-genus and species-family relationships, respectively; where G represents the number of genera, F the number of families and S the number of species. One-way ANOVA was used to test the variation of G/S and F/S ratios among sampling sites.
Spatial similarity (beta diversity) of plant diversity between sampling sites was assessed qualitatively using the Jaccard index using species presence/absence data per site, and quantitatively by Bray-Curtis distance using data of average species densities per site. A clustering analysis (Ward’s method) was applied to the results of the similarity analysis in order to distinguish homogeneous groups of sampling sites “sandy habitats” according to their composition and abundance of psammophyte species.
2.5 Species morpho-funtional traits and chorological features of plant communities
The psammophytic and other plant communities in each sampling site were characterized using several aspects related to the biology, ecology and biogeography of plant species. Using various sources as detailed here below, Raunkiaer lifeform, morphological, dispersal, Noy-Meir, and chorological types were assigned and defined for each plant species.
Plant life forms were first defined by Raunkiaer (1904), then the concept of biological spectrum was introduced by Raunkiaer (1907). The latter helps to determine the proportions of each life forms in the flora of a given site. Thus, to establish the biological spectra by site, life forms of plant communities in sandy habitats were determined based on the work of Quézel (1965). The determination of the morphological types, i.e. annual vs. perennial species, was based on the work of Quézel and Santa (1962; 1963) and Ozenda (2004).
The plants present several dispersal strategies following the types of diaspores. According to the agent involved in the dissemination of diaspores van der Pijl (1982) classified plants into five main categories namely anemochore, autochore, barochore, hydrochore and zoochore. Plants of sandy habitats were classified following this classification where the types of plant dispersal were determined based on Quézel and Santa (1962; 1963), Jauffret (2001) and Bouallala et al. (2020).
Based on plant adaptation strategy against drought, Evenari et al. (1975) and Noy-Meir (1973) classified plants into two categories: arido-passive plants which are defined as species which do not exhibit any photosynthetic activity during the dry period, and arido-active plants which exhibit photosynthetic activity, even reduced, during the dry period. For the classification of plants in sandy habitats of the Sahara Desert according to these categories, previous studies were used to determine the category of each species (Quézel and Santa, 1962; 1963; Ozenda, 2004; Jauffret, 2001; Bradai et al., 2015a; Bouallala et al., 2020).
According to the phytogeographic classification of Quézel (1978), the study area belongs to the Holarctic realm, Mesogean sub-realm, Saharo-Arabian region, Saharan sub-region, and the northern Sahara domain. The determination of the chorological types of plant species encountered in sandy habitats was based on the study of Quézel (1965).
The appreciation of the abundance for plant species identified in sandy soils and dunes of the Sahara Desert was defined based on the works of Quézel and Santa (1962; 1963) on the Algerian flora. Consequently, each plant species was classified according to its degree of rarity in Algeria, which includes these categories (from widespread to extremely rare): particularly widespread (CCC), very common (CC), common (C), fairly common (AC), quite rare (AR), rare (R), very rare (RR), and extremely rare (RRR).
3 Results
3.1 Plant species composition, richness and structure
In general, in the sampled sandy habitats from the Northern Sahara Desert of Algeria we registered 29 plant species belonging to 27 genera and 18 families with a dominance of Brassicaceae (13.79%) and Poaceae (13.79%) (Table 1). During sampling, a total of 1244 individuals/100sqm were identified with a maximum density observed in site 3 (321 individuals/100sqm) followed by site 1 (262 individuals/100sqm) and a minimum in site 7 (80 individuals/100sqm). Among the species encountered, Stipagrostis pungens and Stipagrostis acutiflora showed the highest densities with a total of 426 and 402 individuals/100sqm, respectively. Stipagrostis pungens was dominant in sites 1 and 2 with mean densities of 15.1 ± 6.6 and 14.9 ± 22.5 individuals/relevé, respectively; the minimum was noted in site 3 with (0.4 ± 0.7 individuals/relevé). Stipagrostis acutiflora dominated at sites 3, 5 and 6 with 18.7 ± 7.3, 10.4 ± 6.7 and 11.1 ± 10.7 individuals/relevé, respectively. The singletons i.e. plants recorded with one individual in the entire study were Stipagrostis obtusa, Zilla macroptera, Pituranthos scoparius, Colocynthis vulgaris, Plantago ciliata and Fagonia glutinosa; most of these species occurred mainly in site 1 (Table 1).
| Sampling sites | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Family | Species | Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | Site 7 | Total |
| Poaceae (13.79%) | Stipagrostis pungens Desf. | 15.1 ± 6.6 | 14.9 ± 22.5 | 0.4 ± 0.7 | 6.5 ± 2.7 | 2.9 ± 3.6 | 4.3 ± 4.4 | 5.2 ± 3.9 | 426 |
| Stipagrostis acutiflora Trin. & Rupr. | — | — | 18.7 ± 7.3 | — | 10.4 ± 6.7 | 11.1 ± 10.7 | — | 402 | |
| Stipagrostis obtusa Del. | 0.1 ± 0.3 | — | — | — | — | — | — | 1 | |
| Danthonia forskalii (Vahl) R.Br. | 0.3 ± 0.9 | — | — | — | — | — | — | 3 | |
| Brassicaceae (13.79%) | Oudneya africana R.Br. | — | — | 3 ± 2.9 | 1.5 ± 2 | 2.3 ± 2.7 | — | — | 68 |
| Zilla macroptera (Coss.) Maire | 0.1 ± 0.3 | — | — | — | — | — | — | 1 | |
| Savignya longistyla (Boiss. & Reut.) Maire | — | — | 0.2 ± 0.6 | — | — | — | — | 2 | |
| Malcolmia aegyptiaca Spr. | — | — | 5.7 ± 10.7 | — | — | — | — | 57 | |
| Ephedraceae (3.45%) | Ephedra alata DC. | — | — | — | 2.7 ± 2.6 | 0.1 ± 0.3 | 1.4 ± 1.6 | 1.4 ± 1 | 55 |
| Chenopodiaceae (10.35%) | Cornulaca monacantha Del. | — | — | — | — | 0.1 ± 0.3 | 1.6 ± 2.4 | — | 17 |
| Anabasis articulata (Fordk) Moq. | — | — | — | — | 0.1 ± 0.3 | 0.6 ± 1.3 | 0.1 ± 0.3 | 8 | |
| Salsola vermiculata L. | 0.3 ± 0.7 | 0.7 ± 1.3 | — | — | — | — | — | 7 | |
| Fabaceae (3.45%) | Retama retam Webb | 0.7 ± 1.1 | 0.5 ± 0.9 | — | — | — | — | 0.3 ± 0.7 | 13 |
| Euphorbiaceae (3.45%) | Euphorbia guyoniana Boiss. & Reut. | 0.4 ± 1 | — | 0.2 ± 0.4 | 0.2 ± 0.4 | 0.1 ± 0.3 | — | 1 ± 1.7 | 18 |
| Asteraceae (6.89%) | Rhantherium suaveolens (Desf.) Q. & S. | 5.1 ± 3.7 | — | — | — | — | — | — | 51 |
| Pulicaria crispa (Forsk.) Benth. & Hook | 0.2 ± 0.6 | — | — | — | — | — | — | 2 | |
| Scrophulariaceae (6.89%) | Antirrhinum ramosissimum Coss. & DR. | 0.4 ± 0.7 | — | — | — | — | — | — | 4 |
| Linaria aegyptiaca (L.) Dum. Cours. | 1.7 ± 2.6 | — | — | — | — | — | — | 17 | |
| Boraginaceae (3.45%) | Moltkia callosa (Forsk.) Maire | — | — | 3.7 ± 4.1 | 0.2 ± 0.4 | 0.2 ± 0.4 | 1 ± 2 | — | 51 |
| Apiaceae (3.45%) | Pituranthos scoparius (C. & D.) Benth. & Hook. | 0.1 ± 0.3 | — | — | — | — | — | — | 1 |
| Asclepiadaceae (3.45%) | Pergularia tomentosa L. | 0.1 ± 0.3 | 0.4 ± 0.7 | — | — | — | — | — | 3 |
| Resedaceae (3.45%) | Randonia africana Coss. | 0.1 ± 0.3 | 2.7 ± 4.4 | — | — | — | — | — | 16 |
| Geraniaceae (6.89%) | Monsonia nivea (Dec.) Webb | 0.4 ± 1.3 | — | — | — | — | — | — | 4 |
| Erodium glaucophyllum L’Her. | — | — | 0.2 ± 0.6 | — | — | — | — | 2 | |
| Cistaceae (3.45%) | Helianthemum lippii (L.) Dum. Cours. | 1 ± 3.2 | — | — | — | — | — | — | 10 |
| Cucurbitaceae (3.45%) | Colocynthis vulgaris (L.) Schrad | 0.1 ± 0.3 | — | — | — | — | — | — | 1 |
| Plantaginaceae (3.45%) | Plantago ciliata Desf. | — | 0.2 ± 0.4 | — | — | — | — | — | 1 |
| Zygophyllaceae (3.45%) | Fagonia glutinosa Del. | — | 0.2 ± 0.4 | — | — | — | — | — | 1 |
| Cyperaceae (3.45%) | Cyperus conglomeratus (Rottb.) | — | — | — | — | — | — | 0.2 ± 0.7 | 2 |
| Total density | 262 | 108 | 321 | 111 | 162 | 200 | 80 | 1244 | |
3.2 Taxonomic structures
Overall, the ratio of generic richness to species richness (G/S) in the sandy habitats of the Sahara Desert of Algeria was 0.94 ± 0.13, whereas the ratio of family richness to species richness (F/S) was 0.91 ± 0.15 (Fig. 2). Values of both ratios were high in all sampling sites; it totaled 1.00 in sites 2, 4 and 7, while the lowest value was recorded in site 6 (F/S = 0.77 ± 0.13). According to one-way ANOVA, the values of both ratios varied significantly among the sampling sites, where the F/S ratio (F(6, 63) = 6.86, P < 0.001) revealed slightly higher variability compared to the G/S ratio (F(6, 63) = 6.20, P < 0.001).
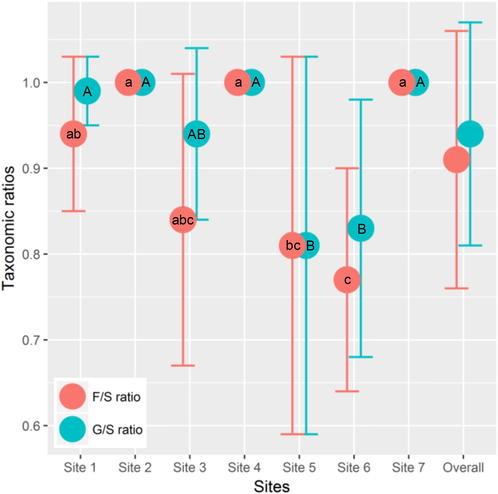
- Ratios of generic richness to species richness (G/S) and of family richness to species richness (F/S) for different sampling sites in sandy habitats of the Algerian Sahara Desert. The same letters associated with the means (solid colored circles) of F/S (lowercase letters) and G/S (uppercase letters) ratios are significantly not different (P > 0.05) among sampling sites following the Tukey’s HSD post hoc test. Vertical error bars are standard deviations.
The linearized power functions assessing the relationships between species richness and richness of higher taxa (i.e. genus and family) are expressed in Fig. 3. The exponents of a in the power model “ln(G or F) = a × ln(S)” exhibited little variation in both genus-species and family-species relationships. According to scores of the goodness-of-fit of the models expressed as R2, the linearized model of the family-species relationships displayed lower stability in comparison to the genus-species relationship. In all sampling sites, both family-species and genus-species relationships were positive and statistically significant (linear regressions: P < 0.01).
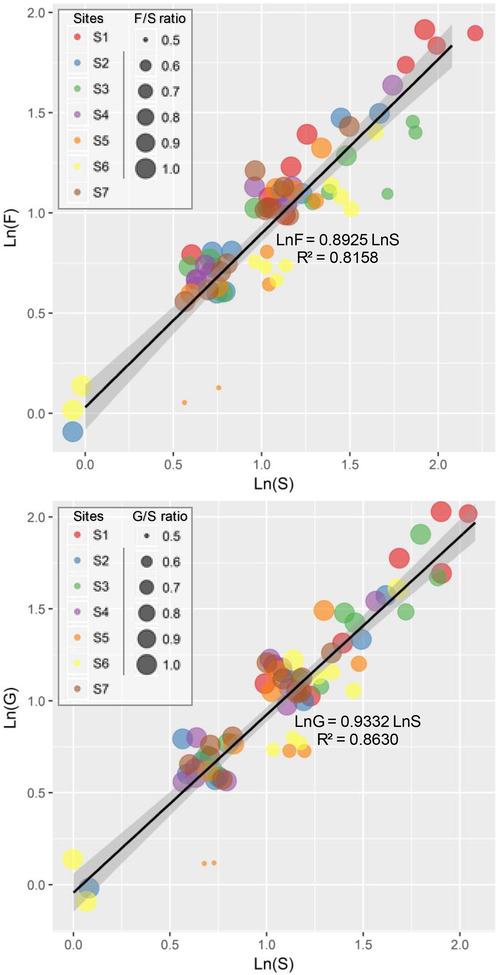
- Relationships between species richness (S) and generic (G)/family (F) richness for different sampling sites in sandy habitats of the Algerian Sahara Desert. The color of points is mapped to sampling sites, and size values are set to F/S (top plot) and G/S (bottom plot) taxonomic ratios. The solid black lines represent a simple linear regression with 95% confidence regions in light gray. The R-squared is a goodness-of-fit measure for the linear regression model.
3.3 Relationships between taxonomic structures and vegetation cover
Pearson’s correlation tests showed that the variations in the number of families, genera and species were positively associated to vegetation cover measured at the level of sampling plots (Fig. 4). While the number of all taxonomic levels was deemed to be positively correlated to plant cover, the number of species (r = 0.51, P < 0.001) then genera (r = 0.49, P < 0.001) showed slightly higher increase with the increase in vegetation cover in comparison to the number of families (r = 0.43, P < 0.001).
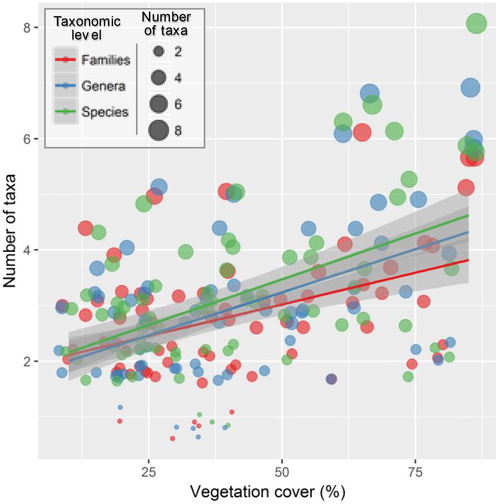
- Relationships vegetation cover and the richness of species, genera and families of psammophytic communities sampled on sand dunes and sandy soils of the northern Sahara Desert. The solid black lines represent a linear regression with 95% confidence regions in light gray.
3.4 Plant diversity
The species richness in sampling sites fluctuated between 5 and 17 species; it varied according to the landscapes. Overall, the floristic diversity of sandy habitats was very low. Species richness ranged between poor in site 1 (S = 17 species) or very poor (<10 species) in the rest of the sampling sites where species richness varied between 5 and 8 (Table 2). The biodiversity measurement revealed that site 1 had the highest value in the Shannon index (H = 1.49) and that site 2 had the lowest value (H = 0.86). In sites 3–7, Shannon index values varied between 1.12 and 1.30. The most balanced plant communities were noted in sites 4 and 6 (E = 0.72), followed by sites 3 and 7 (E = 0.63–0.64). The lowest value of evenness was recorded in site 2. Due to the presence of many rare species, site 1 showed relatively low evenness although it had the highest values of species richness and Shannon index.
| Sampling sites | |||||||
|---|---|---|---|---|---|---|---|
| Diversity parameters | Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | Site 7 |
| Species richness # | 17 ** | 7 * | 8 * | 5 * | 8 * | 6 * | 6 * |
| Shannon index | 1.49 | 0.86 | 1.24 | 1.16 | 1.12 | 1.30 | 1.12 |
| Maximum Shannon index | 2.83 | 1.94 | 1.94 | 1.60 | 2.07 | 1.79 | 1.79 |
| Evenness index | 0.52 | 0.44 | 0.64 | 0.72 | 0.53 | 0.72 | 0.63 |
(#: state of the flora, **: species-poor flora, *: very species-poor flora).
3.5 Analysis of similarity between sites
The Venn diagram shown in Fig. 5 represents the distribution of species between the study sites. The diagram showed that there is a single species (Stipagrostis pungens) omnipresent and common among all the sites studied. Most of shared species were present in site 1. While the sites 4–6 had no exclusive species (i.e. all their species were recorded in other sites), eleven species occurred exclusively in site 1, site 3 had three, site 2 contained two and site 7 had one exclusive species (Fig. 5). These species were Colocynthis vulgaris, Pituranthos scoparius, Stipagrostis obtuse, Zilla macroptera, Pulicaria crispa, Danthonia forskalii, Antirrhinum ramosissimum, Monsonia nivea, Linaria aegyptiaca, Helianthemum lippii, and Rhantherium suaveolens for site 1; Erodium glaucophyllum, Malcolmia aegyptiaca, and Savignya longistyla for site 3; Fagonia glutinosa, and Plantago ciliata for site 2 and Cyperus conglomeratus for site 7 (Table 1).
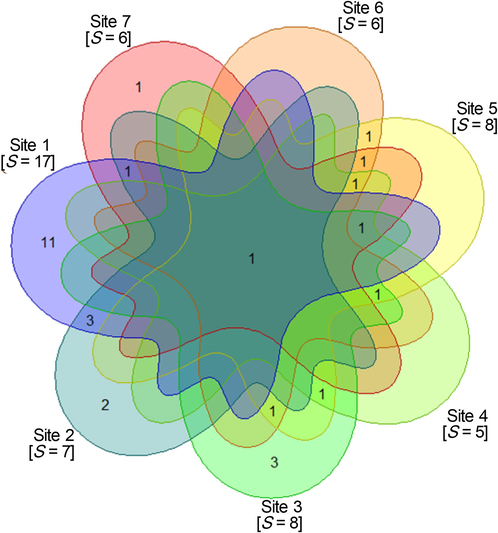
- Seven-set Venn diagram displaying plant species richness (S) recorded at various sites of sand dunes at the Sahara Desert of Algeria. Values of S reported between square brackets represent the total number of species in each sampling site, whereas numbers within the diagram are species shared exclusively between the corresponding sites.
The qualitative analysis of plant community similarity between sites using Jaccard index showed the highest similarity scores between sites 5 and 6 (similarity = 75%) and sites 4 and 5 (similarity = 62.5%). These three sites along with sites 3 and 7 were clustered as a homogenous group which had high similarities of plant composition (Fig. 6). Besides, sites 1 and 2 were clustered a unique group (similarity = 26.3%). These two sites exhibited low similarities (<18%) with sites composing the first cluster. For the quantitative similarity computed using Bray-Curtis distance based on plant abundance data, sites 5–6 (similarity = 85%), sites 1–2 (similarity = 64.4%) and sites 4–7 (similarity = 58.2%) represented the highest similarity values. All other similarities were <50%, except for the similarity between sites 3–5 (similarity = 53.6%). The clustering analysis revealed that sites 1, 2, 4 and 7 had quantitatively similar plant composition, whereas sites 3, 5 and 6 were also homogenous. Intergroup similarities between sites composing these two groups were low.
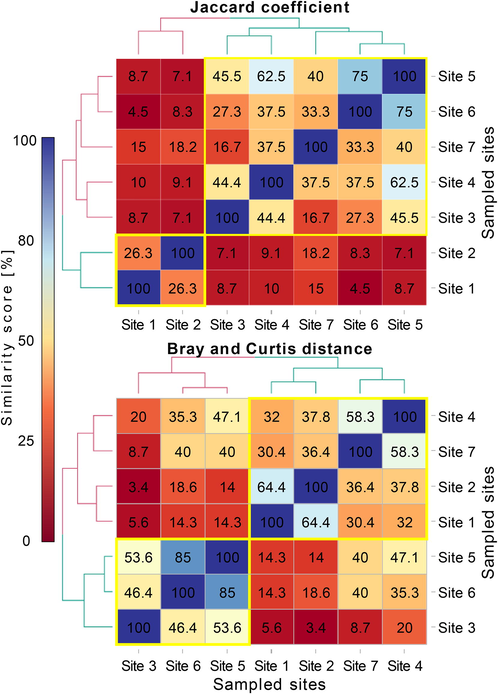
- Heatmaps, with dendrograms (side colored plots) of hierarchical clustering analysis (Ward’s method), displaying the matrix of plant similarities between sampling sites of sandy habitats in the Sahara Desert of Algeria. Similarity was displayed in color intensity proportionally to similarity scores: Jaccard’ coefficient (upped plot) and Bray and Curtis distance (bottom plot). Yellow rectangles within the heatmaps matrices represent homogeneous clusters of sites based on plant composition.
3.6 Plant functional traits
With the exception of sites 3 and 7, chamaephytic species had the highest values in the biological spectra of lifeforms in sites 1, 2, 4, 5 and 6 with 10, 3, 3, 5 and 4 species, respectively (Table 3, Fig. 7). Hemicryptophytes, with 3 species, had slightly higher values than chamaephytes in sites 3 and 7. Therophytes were only observed in sites 1, 2 and 3 with respectively 2, 2 and 3 species. Phanerophytes were represented by one species (Retama retam), which occurred at sites 1, 2 and 7. As morphological types were concerned, the flora was composed of 7 annuals and 22 perennial plants. While the annual species were completely absent in sites 4–7, perennials were numerous and well represented in sites 1–3, with a higher value at site 1 (15 species). Basically in almost all sampling sites, the dominant plant dispersal types were anemochores then barochores. The zoochore type was weakly represented, where only three species were recorded at site 1. Regarding the Noy-Meir's classification, arido-active plants dominated in all sampling sites, except site 3, with a good representation of species in site 1 (12 species) and sites 5 and 6 with six species for each (Table 3, Fig. 7).
| Plant species | Ecological and functional traits | ||||||
|---|---|---|---|---|---|---|---|
| Morphological types | Life forms | Chorological types | Abundance appraisal | Dispersal types | Noy-Meir types | Spatial distribution | |
| Stipagrostis pungens | Perennial | Hemi | SA | CC | Anemo | Arido-active | 1–7 |
| Stipagrostis acutiflora | Perennial | Hemi | SA | R | Anemo | Arido-active | 3, 5, 6 |
| Stipagrostis obtusa | Perennial | Hemi | MSA | C | Anemo | Arido-active | 1 |
| Danthonia forskalii | Perennial | Hemi | SA | CC | Anemo | Arido-passive | 1 |
| Oudneya africana | Perennial | Cham | End-S | AR | Baro | Arido-passive | 3–5 |
| Zilla macroptera | Perennial | Cham | SA | C | Zoo | Arido-active | 1 |
| Savignya longistyla | Annual | Ther | SA | C | Baro | Arido-passive | 3 |
| Malcolmia aegyptiaca | Annual | Ther | SA | C | Baro | Arido-passive | 3 |
| Ephedra alata | Perennial | Cham | SA | CC | Anemo | Arido-active | 4–7 |
| Cornulaca monacantha | Perennial | Cham | SA | AC | Baro | Arido-active | 5, 6 |
| Anabasis articulata | Perennial | Cham | SA | C | Anemo | Arido-active | 5–7 |
| Salsola vermiculata | Perennial | Cham | MSA | C | Anemo | Arido-active | 1, 2 |
| Retama retam | Perennial | Phan | SA | C | Baro | Arido-active | 1, 2, 7 |
| Euphorbia guyoniana | Perennial | Hemi | End-S | CC | Baro | Arido-passive | 1, 3–5, 7 |
| Rhantherium suaveolens | Perennial | Cham | SA | R | Zoo | Arido-active | 1 |
| Pulicaria crispa | Perennial | Cham | SA | AC | Anemo | Arido-active | 1 |
| Antirrhinum ramosissimum | Perennial | Cham | SA | C | Baro | Arido-active | 1 |
| Linaria aegyptiaca | Perennial | Cham | MSA | C | Baro | Arido-active | 1 |
| Moltkia callosa | Perennial | Cham | SA | C | Anemo | Arido-active | 3–6 |
| Pituranthos scoparius | Perennial | Cham | MSA | C | Zoo | Arido-active | 1 |
| Pergularia tomentosa | Perennial | Cham | SA | CC | Anemo | Arido-active | 1, 2 |
| Randonia africana | Perennial | Cham | SA | R | Baro | Arido-active | 1, 2 |
| Monsonia nivea | Annual | Ther | SA | CC | Anemo | Arido-passive | 1 |
| Erodium glaucophyllum | Annual | Ther | MSA | C | Anemo | Arido-passive | 3 |
| Helianthemum lippii | Perennial | Cham | MSA | CCC | Baro | Arido-passive | 1 |
| Colocynthis vulgaris | Annual | Ther | MSA | CC | Anemo | Arido-passive | 1 |
| Plantago ciliata | Annual | Ther | SA | CC | Baro | Arido-passive | 2 |
| Fagonia glutinosa | Annual | Ther | SA | CC | Baro | Arido-passive | 2 |
| Cyperus conglomeratus | Perennial | Hemi | SA | C | Anemo | Arido-active | 7 |
Life forms: [Cham: chamaephyte, Hemi: hemicryptophyte, Phan: phanerophyte, Ther: therophyte];
Chorological types (identified according to Quézel and Santa, 1962; 1963) [End-S: endemic to the Sahara Desert, MSA: Mediterranean Saharo-Arabian, SA: Saharo-Arabian];
Abundance appraisal (determined following the classification of Quézel and Santa (1962; 1963): [CCC: Widespread, CC: Very common, C: Common, AC: Fairly common, AR: Quite rare, R: Rare];
Dispersal types: [Anemo: anemochores, Baro: barochores, Zoo: zoochores];
Spatial distribution: figures [1–7] refer to sampling sites where the species occurred.
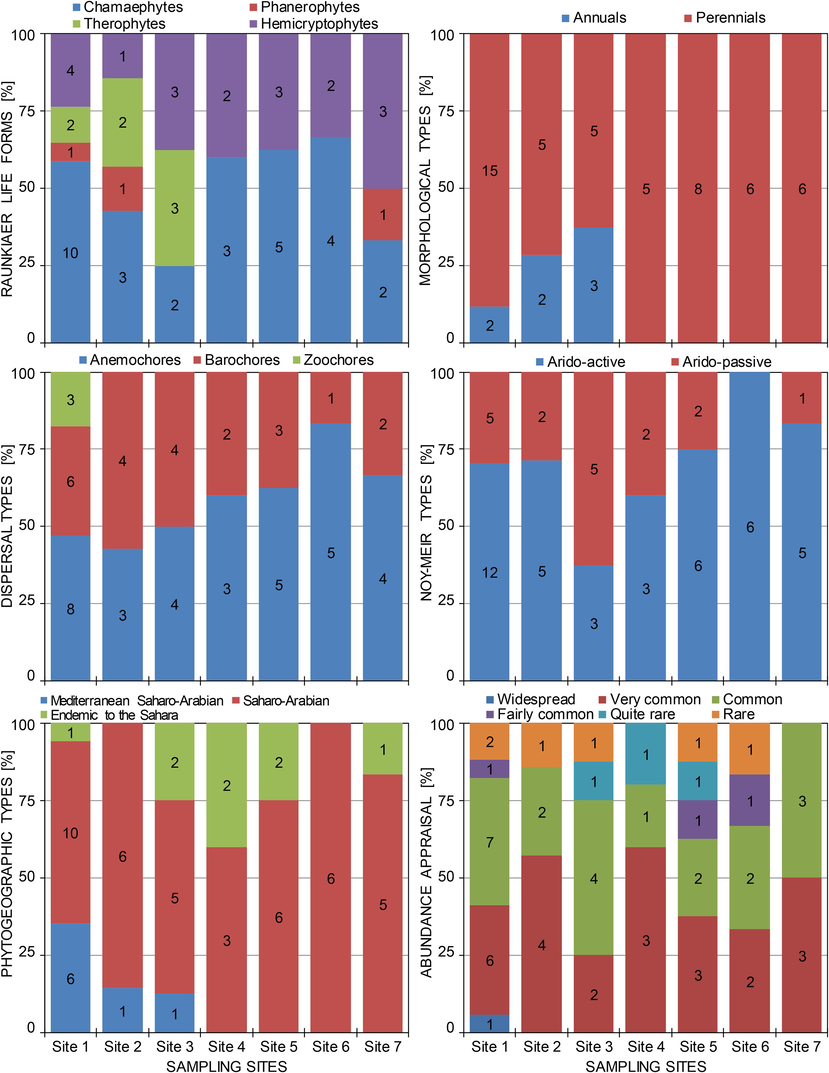
- Distribution of ecological and functional traits (life forms, morphological types, dispersal types, Noy-Meir’s types, chorological types, and abundance appraisal categories) for different sampling sites of sandy soils and dunes in the Sahara Desert of Algeria. Figures within histograms refer to the number of plant species.
3.7 Chorological types
The analysis of phytogeographic (chorological) characteristics of the flora of sandy habitats of the northern Sahara Desert revealed a strong presence of the Saharo-Arabian chorological element in all sampling sites, with the highest level (10 species) recorded in site 1 and lowest level (3 species) noted in site 4 (Table 3, Fig. 7). The Mediterranean-Saharan-Arabian chorological element was observed only at sites 1–3 with 6, 1 and 1 species, respectively. As for the endemism, we noted the presence of the Saharan endemic element at sites 1, 3, 4, 5 and 7 represented by two species - Oudneya africana and Euphorbia guyoniana.
3.8 Appreciation of species abundance
Plant species of psammophytic communities inhabiting sandy soils and dunes of the Sahara Desert of Algeria included six categories of abundance for the flora of Algeria (AC: fairly common, C: common, CC: very common, CCC: particularly widespread, AR: fairy rare, and R: rare) (Table 3, Fig. 7). The particularly widespread category was recorded at site 1 with a single species (Helianthemum lippii). The common and very common categories were observed at all sites with varying rates. The fairly common category was represented by two species (Cornulaca monacantha and Pulicaria crispa), which were identified at sites 1, 5 and 6. The fairly rare category was represented by a single species (Oudneya Africana) recorded at sites 3–5. Whereas the category of rare species was represented by three species (Stipagrostis acutiflora, Rhantherium suaveolens and Randonia Africana), which occurred in sites 1, 2, 3, 5 and 6.
4 Discussion
This study represents a comprehensive analysis of diversity of plant communities in desert sandy environments such as dunes and sandy soils in northern Algerian Sahara Desert. The characterization of plant species diversity and biological traits of vegetation communities in natural habitats is an essential procedure for the preservation of species, communities, habitats and ecosystems (Gamoun, 2013). The characteristic plant families of sandy environments in the study area are Poaceae, Brassicaceae and Chenopodiaceae, these families adapt well to the ecological conditions of most Saharan habitats. This is in agreement with studies of flora and vegetation of arid and hyper-arid regions (Monod, 1992, Chehma, 2005, Bouallala, 2013, Gamoun et al., 2018). Based on plant abundance data, perennial species are the characteristic morphological type of vegetation in sandy soils of the northern Sahara Desert, where Stipagrostis pungens and Stipagrostis acutiflora dominated in terms of plant density with 34% and 32% of the total, respectively. These two species create favorable environmental conditions for the establishment and development of other plants in sandy habitats and also provide an important fodder resource for feeding dromedaries raised in extensive grazing system (Chehma et al., 2008a;b; Bouallala, 2013; Chehma and Huguenin, 2017). In hot deserts, perennial and dominant plants play an important ecological role in the stabilization of degraded sandy soils and the fixation of sand dunes by trapping aeolian sediments(Chenchouni et al., 2019, 2022; Rustamov, 1994). Furthermore, desert regions are often used throughout the year for camel, sheep and goat free grazing and natural vegetation is a valuable feed resource which plays a crucial role in the livestock industry at the regional and country level (Rustamov, 1994). In this vein, livestock grazing has several impacts on plant communities. According to Merdas et al. (2021), grazing in arid steppe rangelands can induce large-scale homogenization in plant communities. However, the response of vegetation to livestock grazing may vary following other covariates including soil types and climates. For instance, vegetation response on sandy and gravelly soils is more diversified and more productive than on limestone and loamy soil, whereas the latter is more adapted to grazing pressure (Gamoun 2013; Gamoun et al., 2012; 2018).
Out of the 18 families, 12 were represented by one species. The species-higher taxon relationships, i.e. relationships between species richness and generic or family richness revealed high scores. This represents a good indicator for a flora with low diversity, which is the key characteristic of desert plant communities. These findings are similar to taxonomic compositions in grazed steppe rangelands of arid and semiarid Algeria (Macheroum et al., 2021; Senoussi et al., 2021). In hot regions with accentuated aridity, most plant families have only one or two genera and most genera includes only one or two species (Kouba et al., 2021). This is in agreement with our findings where the G/S ratio was higher than 81% and F/S ratio > 77%. Species richness at the site scale is poor or even very poor in all studied sites of sandy soils and dunes, which reflects the low diversity of plants in these environments. These findings are comparable to results from previous studies analyzing the vegetation of sandy soils of rangelands in the northern Algerian Sahara Desert (Chehma, 2005) or the desert of Turkmenistan, which occupy > 80% of the country territory, and where very few semi-shrub species dominate and provide most of the phytomass (Rustamov, 1994). Bouallala (2013) indicated moderate flora diversity in sandy soils of arid rangelands in western Algerian Sahara Desert. The differences in plant diversity and taxonomic composition between hot desert regions may be due to different climatic and edaphic conditions of the habitat (Abd El-Ghani et al., 2015; Alyemeni, 2000; El-Sheikh et al., 2021; Pavlik, 1989; Salama et al., 2014).
The prominence of chamaephytes in the studied sandy soils reflects their good adaptation to the aridity of the Saharan climate (Chenchouni, 2012; Bouallala, 2013; Bradai et al., 2015a; Gamoun et al., 2012; 2018). Hemicryptophyes are represented by four species of Poaceae family, one species of Euphorbiaceae and one species of Cyperaceae. The good representativeness of the Poaceae in this study seems to indicate their good adaptation strategy to sandy environments. Bouallala (2013) confirmed that the presence and abundance of hemicryptophytes in sandy soils and dunes are represented by psammophytes, in particular perennials, belonging to Poaceae family. According to Barbero et al. (2001) the abundance of hemicryptophytes in Maghreb countries is related to site-specific soil conditions, in particular soil moisture and organic matter levels. Therophytes (annual species) were poorly represented in the studied sandy habitats and are found in only three sites. This low representation of theophytes is probably linked to the lack of precipitation in the Saharan environment and also to the physicochemical characteristics of the soil that has poor water holding capacity. Several studies in the Sahara Desert have emphasized that the therophytes represent the biological type which depends directly on precipitation amount and rainfall periodicity (Quézel, 1965; Barkoudah and van der Sar, 1982; Monod, 1992). As an evolutionary adaptation strategy, therophytes escape extreme conditions as seeds. Phanerophytes were represented by a single species i.e. Retama retam which was present in three sites only. Generally, phaneophytes are rare in the Sahara and hot deserts (Bouallala, 2013; Gamoun et al., 2018) and are localized in mesic habitats receiving irrigation water such as wadi beds and irrigation channels (Bouallala, 2013; Bradai et al., 2015a; Bouallala et al., 2020). According to Rustamov (1994), annual ephemerous species have very little contribution in structuring plant communities in desert of Turkmenistan.
With the exception of site 2, anemochory is the most represented dispersal plant strategy followed by barochory. On the other hand, the zoochoric species, located at site 1, were less represented with only three species (Zilla macroptera, Rhanterium suaveolens and Pithuranthos scoparius). Wind is an essential driver in the dispersion of the diaspores of the majority of plants in flat landscapes of the Sahara Desert (Bradai et al., 2015a; Bouallala et al., 2020). Barochoric species have special physiological mechanisms to ensure seed dispersal (Bouallala et al., 2020). The presence of animals in Saharan environments ensures dispersal of many plant species, sometimes with remarkable specialization in zoochorous plant-animal interactions (Bradai et al., 2015a).
The Noy-Meir classification shows a good representation of arido-active plants in almost all studied sites. This can be explained by the site-specific edapho-climatic and water conditions of sandy soils and dunes of the Saharan. As most species in sandy soils of the study area are perennials, these arido-active plants are dependent of soil water status. According to Jauffret (2001), arido-active species have water reserves to function and survive under the harsh environmental conditions of the hot deserts. It is worth mentioning that the abundance and diversity of arido-passive plants were low in the study area, which is explained by the fact that the majority of these plants are annuals and depend exclusively on erratic and seasonal rainfall.
The Saharo-Arabian species are the dominant chorological type in all studied sites at the Sahara Desert. With seven species, the Mediterranean-Saharan-Arabian plants, are localized only at sites 1–3. Endemic plants to the Saharan included two species: Euphorbia guyoniana observed in sites 1, 3–5 and 7, and Oudneya africana occurring in sites 3–5. The good representation of the Saharo-Arabian chorological elements in Saharan environments was reported in many studies of the Saharan flora (Salama et al., 2014; Bradai et al., 2015a; Bouallala et al., 2020). Mediterranean-Saharan-Arabian plants find appropriate environmental conditions for their development in the Sahara Desert, probably through compensatory factors, in particular soil water. The endemic elements represent very specialized species with a limited geographic areal.
The appraisal of plant abundance showed a good representation of very common and common plants in all studied sites. Rare species were present at sites 1–3, 5, 6, quite rare species occurred at sites 3–5, fairly common species develop at sites 1, 5 and 6. The herbaceous plant Helianthemum lippii, a unique species in Algeria, was recorded at site 1. In various Saharan habitats, this species forms mycorrhizae with several species of desert truffles (Bradai et al., 2013; 2014), which have an economic importance for desert people (Bradai et al., 2015c). The presence and abundance of plants in the Saharan environment are determined by abiotic factors related to the climate, geomorphology and soil, and biotic interactions represented mainly by anthropogenic factors and interspecific competition (Ozenda, 2004; Chehma, 2005; Bouallala et al., 2020). Sandy soils and dunes are unique habitats in the northern Algerian Sahara Desert that have specific peculiarities offering sustainable resources for human well-being (namely food, firewood, medicinal plants), feeding domestic and wild animals and offering an ecological niche and food resources for other Saharan living beings (Longo-Hammouda et al., 2007; Chehma et al., 2008a; Bouallala, 2013; Bradai et al., 2020). Out of 29 plant species identified in this study, two were endemic to the Sahara Desert, three were rare, two were fairly common and one species was quite rare. These rare species are endangered and at risk of decline or even extinction due to the abusive use and overexploitation by man (uprooting, cutting wood, clearing). In order to preserve this vulnerable and rare flora, specific management measures are required.
5 Conclusion
This work highlighted the importance of studying plant diversity in sandy soils and dunes of the northern Algerian Sahara Desert. Species richness in these Saharan habitats reached 29 plants represented mainly by perennial and arido-active species. Although the flora was qualified as poor in terms of species richness compared to other floras in the world, in terms of hot deserts, it is of high value. This permanent vegetation of sand habitats provides food for various Saharan living beings throughout the year. Arido-passive plants, represented by therophytes, are less abundant in sandy habitats, but these plants are an important resource for free grazing livestock animals, mainly dromedary, as well as for wild herbivores during periods that follow rainfall events. The characterization of plant lifeforms confirms the good adaptation of chamaephytes to ecological conditions of arid and hyperarid hot environments through the development of specific strategies specific to this category of plants. The Saharan-Arabian chorological element is a good indicator of the Saharan climate, whereas the presence of the Saharan endemic element translates the specificity of certain plants that inhabit and prefer the Saharan environment. The type of dispersal that dominates in dunes and sandy habitats is anemochory , in which case the wind is the main dispersal factor of diaspores of the majority of plants. Abundance appraisal is an essential aspect in decision-making related to the protection of species and their habitats, especially in sandy habitats which are very fragile, unstable and threatened by various degradation factors. Under the current environmental and socioeconomic conditions of a changing world, it is highly recommended to update the status of species in terms of rarity and conservation.
6 Availability of data
The datasets used and/or analyzed during the current study are available from the corresponding author on a reasonable request.
CRediT authorship contribution statement
Marouane Azizi: Methodology, Investigation. Haroun Chenchouni: Formal analysis, Visualization, Writing – review & editing. Mohammed El Hafedh Belarouci: Investigation, Data curation. Lyès Bradai: Investigation, Writing – original draft. M'hammed Bouallala: Conceptualization, Methodology, Data curation, Investigation, Resources, Writing – original draft.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Habitat diversity and floristic analysis of Wadi El-Natrun Depression, Western Desert, Egypt. Phytol. Balc.. 2015;21(3):351-366.
- [Google Scholar]
- Abdelkrim, H., 2006. Note méthodologique pour la mise en place d’un système de suivi de la biodiversité. Rapport scientifique. Projet PNUD 0034575 “Préservation et utilisation de la diversité biologique d’intérêt mondial dans les parcs nationaux de l’Ahaggar et du Tassili”. 32p.
- Ecological studies on sand dunes vegetation in AI-Kharj region, Saudi Arabia. Saudi J. Biol. Sci.. 2000;7:64-87.
- [Google Scholar]
- Signification biogéographique et biodiversité des forêts du bassin Méditerranéen. Bocconea. 2001;13:11-25.
- [Google Scholar]
- L’Acacia raddianadans la région de Béni Abbès (Algérie) Bull. Soc. Hist. Nat. Afr. N.. 1982;70:79-121.
- [Google Scholar]
- Essai de monographie du Cupressus dupreziana A. Camus, cyprès endémique du Tassili des Ajjer (Sahara Central) Bull. Soc. Hist. Nat. Afr. N.. 1970;61:95-178.
- [Google Scholar]
- Vegetation associations in the Great Western erg and the Saoura Valley, Algeria. Phytocoenologia. 2001;31(3):311-324.
- [CrossRef] [Google Scholar]
- Vegetation associations in the Ougarta Mountains and dayas of the Guir Hamada, Algerian Sahara. J. Arid Environ.. 2003;54(4):739-753.
- [CrossRef] [Google Scholar]
- Antimicrobial activity of xerophytic plant (Cotula cinerea Delile) extracts against some pathogenic bacteria and fungi. Jordan J. Biol. Sci.. 2013;6(4):266-271.
- [Google Scholar]
- Effects of sand encroachment on vegetation diversity in the Sahara Desert. In: Chenchouni H., Chaminé H.I., Khan M.F., Merkel B.J., Zhang Z., Li P., Kallel A., Khélifi N., eds. New Prospects in Environmental Geosciences and Hydrogeosciences. Cham: Springer; 2022.
- [Google Scholar]
- Évaluation de la valeur nutritive de quelques plantes herbacées broutées par le dromadaire dans le Sahara nord occidental algérien. Lebanese Sci. J.. 2013;14(1):33-39.
- [Google Scholar]
- Vegetation traits are accurate indicators of how do plants beat the heat in drylands: Diversity and functional traits of vegetation associated with water towers in the Sahara Desert. Ecol. Indic.. 2020;114:106364.
- [CrossRef] [Google Scholar]
- Etude floristique et nutritive spatio-temporelle des parcours camelins du Sahara Occidental Algérien: Cas des régions de Béchar et Tindouf. Algeria: University of Ouargla; 2013. PhD thesis
- Bouallala, M., Chehma, A., Bensetti, M., 2011. Chemical composition variability of main grazed plant by the dromedary in the South western of Algeria. Livestock Research for Rural Development, 23(5), 107. www.lrrd.org/lrrd23/5/Boua23107.htm.
- Etude mycologique et bio-écologique de la truffe blanche du désert (Tirmania nivea Desf. Trappe 1971) dans la région de Oued M’ya (Ouargla, Sahara algérien) Rev. Bioressources. 2013;3:6-14.
- [Google Scholar]
- Desert truffles of the North Algerian Sahara: diversity and bioecology. Emir. J. Food Agric.. 2014;26(5):425-435.
- [CrossRef] [Google Scholar]
- An appraisal of eremophyte diversity and plant traits in a rocky desert of the Sahara. Folia Geobot.. 2015;50(3):239-252.
- [CrossRef] [Google Scholar]
- Effects of climate on the productivity of desert truffles beneath hyper-arid conditions. Int. J. Biometeorol.. 2015;59(7):907-915.
- [CrossRef] [Google Scholar]
- Diversité et utilisation des plantes médicinales cultivées dans le Sud de Oued Righ. Afr. Rev. Sci. Tech. Dev.. 2020;5(1):61-70.
- [Google Scholar]
- Le « Drinn », Stipagrostis pungens. Caractéristiques floristiques et modes d’usage. Rev. BioRessources. 2017;7:59-66.
- [Google Scholar]
- Chehma, A., 2005. Etude floristique et nutritive spatio-temporelle des parcours camelins du Sahara septentrional algérien. Cas des régions de Ouargla et Ghardaia. Thèse de Doctorat. Université d'Annaba, 178 p.
- Ethnomycological survey of traditional usage and indigenous knowledge on desert truffles among the native Sahara Desert people of Algeria. Journal of Ethnopharmacology. 2015;162:31-38.
- [CrossRef] [Google Scholar]
- Productivité de la phytomasse éphémère des parcours camelins du Sahara septentrional Algérien. Fourrages. 2008;194:253-256.
- [Google Scholar]
- Productivité fourragère et capacité de charge des parcours camelins du Sahara septentrional Algérien. Sécheresse. 2008;19(2):115-121.
- [Google Scholar]
- Diversité floristique d’un lac du Bas-Sahara algérien. Acta Bot. Malac.. 2012;37:33-44.
- [Google Scholar]
- Edaphic factors controlling the distribution of inland halophytes in an ephemeral salt lake “Sabkha ecosystem” at North African semi-arid lands. Sci. Total Environ.. 2017;575:660-671.
- [CrossRef] [Google Scholar]
- New Prospects in Environmental Geosciences and Hydrogeosciences. Cham: Springer; 2022.
- Exploring the Nexus of Geoecology, Geography, Geoarcheology and Geotourism: Advances and Applications for Sustainable Development in Environmental Sciences and Agroforestry Research. Cham, Switzerland: Springer; 2019.
- Hommes et plantes au Sahara, comment hommes et plantes se sont adaptés à des conditions écologiques difficiles. Sahara (Segrate). 2012;23:172-182.
- [Google Scholar]
- Daget, P., Poissonet, J., 1991. Prairies permanentes et pâturages. Méthodes d’étude. Montpellier, France, Institut de Botanique. 354 p.
- Ecology of inland sand dunes “nafuds” as a hyper-arid habitat, Saudi Arabia: floristic and plant associations diversity. Saudi J. Biol. Sci.. 2021;28(3):1503-1513.
- [CrossRef] [Google Scholar]
- Adaptive mechanisms in desert plants. In: Vernberg F.J., ed. Physiological Adaptation to the Environment. Cambridge University Press; 1975. p. :111-129.
- [Google Scholar]
- Analysing taxonomic structures and local ecological processes in temperate forests in North Eastern China. BMC Ecol.. 2017;17(1):1-11.
- [CrossRef] [Google Scholar]
- Diversity of desert rangelands of Tunisia. Plant Divers.. 2018;40(5):217-225.
- [CrossRef] [Google Scholar]
- Vegetation change in variable rangeland environments: the relative contribution of drought and soil type in arid rangelands. Ekológia. 2013;32:148-157.
- [CrossRef] [Google Scholar]
- Impact of grazing on the floristic diversity of arid rangelands in South Tunisia. Rev. Ecol-Terre Vie. 2012;67:271-282.
- [Google Scholar]
- Traditional medicine in Central Sahara: Pharmacopoeia of Tassili N’ajjer. J. Ethnopharmacol.. 2006;105(3):358-367.
- [CrossRef] [Google Scholar]
- Validation et comparaison de divers indicateurs des changements à long terme dans les écosystèmes méditerranéens arides: Application au suivi de la désertification dans le Sud tunisien. France: Univ. Aix-Marseille 3; 2001. Doctoral thesis
- Plant community composition and structure under short-term grazing exclusion in steppic arid rangelands. Ecol. Indic.. 2021;120:106910.
- [CrossRef] [Google Scholar]
- Aspects nutritionnels des pâturages les plus appréciés par Camelus dromedarus en Algérie. Cah. Agric.. 2007;16:477-483.
- [CrossRef] [Google Scholar]
- Médail, F., Quézel, P., 2018. Biogéographie de la flore du Sahara. Une biodiversité en situation extrême, IRD Éditions et Éditions des Conservatoire et jardin botaniques de Genève, Marseille, 366 p.
- Livestock grazing-induced large-scale biotic homogenization in arid Mediterranean steppe rangelands. Land Degrad. Dev. 2021
- [Google Scholar]
- Monod, T., 1992. Du désert. Sécheresse 3(1), 7–24.
- Environmental drivers of taxonomic and phylogenetic diversity patterns of plant communities in semi-arid steppe rangelands of North Africa. Ecol. Indic.. 2021;132:108279.
- [CrossRef] [Google Scholar]
- New insights for the production of medicinal plant materials: ex vitro and in vitro propagation of valuable Lamiaceae species from northern Africa. Curr. Plant Biol.. 2021;27C:100216.
- [CrossRef] [Google Scholar]
- Rehabilitation of Degraded Rangeland in Drylands by Prickly Pear (Opuntia ficus-indica L.) Plantations: effect on soil and spontaneous vegetation. Ecol. Balkan.. 2013;5(2):63-83.
- [Google Scholar]
- Floristic composition and analysis of spontaneous vegetation of Sabkha Djendli in North-east Algeria. Plant Biosyst.. 2016;150(3):396-403.
- [CrossRef] [Google Scholar]
- Diversity and functional traits of spontaneous plant species in Algerian rangelands rehabilitated with prickly pear (Opuntia ficus-indica L.) plantations. Turk. J. Bot.. 2018;42:448-461.
- [CrossRef] [Google Scholar]
- Desert ecosystems: environment and producers. Annu. Rev. Ecol. Evol. Syst.. 1973;4(1):25-51.
- [CrossRef] [Google Scholar]
- Ozenda, P., 2004. Flore et végétation du Sahara 3 édition. Ed. CNRS Paris. 662 p.
- Phytogeography of sand dunes in the Great Basin and Mojave Deserts. J. Biogeogr.. 1989;16:227-238.
- [CrossRef] [Google Scholar]
- La végétation du Sahara du Tchad à la Mauritanie. Stuttgart: Gustav Verlag; 1965. p. :333.
- Analysis of the flora of Mediterranean and Saharan Africa. Ann. Mo. Bot. Gard.. 1978;65(2):479-534.
- [Google Scholar]
- Quézel, P., Santa, S., 1962. Nouvelle flore de l'Algérie et des régions désertiques méridionales. Vol. 1. Ed. CNRS Paris.
- Quézel, P., Santa, S., 1963. Nouvelle flore de l'Algérie et des régions désertiques méridionales. Vol. 2. Ed. CNRS Paris.
- Biological types with reference to the adaptation of plants to survive the unfavourable season. Raunkiaer. 1904;1934:1-2.
- [Google Scholar]
- The life-forms of plants and their bearing on geography. Raunkiaer. 1907;1934:2-104.
- [Google Scholar]
- Rustamov, I.G., 1994. Vegetation of the Deserts of Turkmenistan. In: Fet, V., Atamuradov, K.I. (eds) Biogeography and Ecology of Turkmenistan. Monographiae Biologicae, vol 72. Springer, Dordrecht, pp. 77-104. https://doi.org/10.1007/978-94-011-1116-4_6.
- Variations in vegetation structure, species dominance and plant communities in South of the Eastern Desert-Egypt. Not. Sci. Biol.. 2014;6(1):41-58.
- [Google Scholar]
- Botanical composition and aroma compounds of semi-arid pastures in Algeria. Grass Forage Sci.. 2021;76(2):282-299.
- [CrossRef] [Google Scholar]
- van der Pijl, L., 1982. Principles of dispersal in higher plants. Springer, Berlin. https://doi.org/10.1007/978-3-642-87925-8.
Appendix A
Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jksus.2021.101656.
Appendix A
Supplementary data
The following are the Supplementary data to this article:




