

Translate this page into:
Analyzing the impact of phosphorous and nitrogen on Castanopsis sclerophylla early growth stages
⁎Corresponding authors. 02144@zjhu.edu.cn (Nengzhi Tan), khan_eco@scbg.ac.cn (Muhammmad Sadiq Khan)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Plant growth elements, particularly nitrogen (N) and phosphorus (P) are vital for their growth and development, particularly for understory vegetation and their excess limits the net productivity of terrestrial ecosystems. This study focuses on the understory vegetation responds and adaptation to key essential nutrients under changing climate scenarios in subtropical evergreen broad-leaved forests, still needs research attention. this, we set up an experiment taking four treatments in a 50-year-old Castanopsis sclerophylla secondary forest under (a) control (CK), (b) N, (c) P, and (d) combined N and P addition, applied to natural forest regeneration seedlings of C. sclerophylla attained similar growth parameters of diameter of 3 cm and 10 cm height. In addition, carbon, N, P, and non-structural carbohydrates (NSC) were determined through the anthrone colorimetric approach in different parts of seedlings. Results show that the combined N + P application enhanced the N and P by 14.48 %−140.55 % in the seedlings in both dry and wet seasons, respectively. However, during wet season, the content of NSC in the plant leaves significantly exceeded under P addition. Remarkably, CK showed increased P in the growing season but lower during the dry season. Furthermore, the root starch content of seedlings showed a significant increase under the application of N and P compared to combined N + P, ranging between 45.60 % and 58.70 %. Overall, the plant growth is attributed to N and P intake. The nutrient addition and seasonal variations have a coupled effect on seedling growth as proved in the in the natural open forest experiment. The study outcomes emphasize that the alterations in NSC allocation in the roots and leaves of C. sclerophylla seedlings under N + P addition could enhance their adaptation to future global climate changes, drought conditions, and high N concentrations.
Keywords
Castanopsis sclerophylla
Non-structural carbohydrates
Subtropical broad leaved
Secondary forest
Seedling

1 Introduction
Plant essential elements nitrogen (N) and phosphorus (P) are indispensable for their growth and development, playing pivotal roles in sustaining the terrestrial ecosystem's net primary productivity (Dickman et al., 2015; Zheng et al., 2016; Feng and Zhu, 2021). However, their natural distribution is highly uneven, limiting ecosystem productivity in regions with deficient N and P (Güsewell, 2004; Elser et al., 2007). With the advent of industrialization, N deposition has escalated, leading to a progressive saturation of forest ecosystems with N (IPCC et al., 2018). On the contrary, P deposition has exhibited relatively low levels compared to N, thereby exacerbating the limitation imposed by phosphorus in ecological contexts (Verhoeven et al., 1996). Particularly in tropical, subtropical, and geologically stable areas, P loss over time due to soil weathering has led to exceedingly low soil P levels, profoundly influencing ecosystem structure and productivity (Dickman et al., 2015; Zheng et al., 2016; Feng and Zhu, 2021; Li et al., 2024b).
Typically, plant photosynthesis and growth are hampered by drought, thereby impacting biomass allocation and growth rates. However, studies about the intricate interplay among N, P, and water on biomass are still rare (O'Brien et al., 2014; Mo et al., 2021). Photosynthesis is the primary avenue for plants to synthesize carbon elements for biomass accumulation. Studies elucidate that a significant portion of leaf N is directed towards diverse photosynthetic mechanisms (Guan and Wen, 2011; Liu et al., 2019), influencing carboxylation capacity and electron transfer rates (Guan and Wen, 2011). In the short term, N deposition can elevate leaf nitrogen content, foster the synthesis of photosynthetic pigments, enhance nutrient accumulation, and stimulate plant growth, thereby augmenting biomass accumulation (Feng and Zhu, 2021; Li et al., 2021). Zak et al. (2000) studied the impact of varying N levels for tree biomass, and outcomes revealed a notable enhancement in tree biomass concomitant with increased N concentration, marking a remarkable 200 % increase compared to low N conditions. Moreover, the augmentation of above-ground growth paralleled the increase in N application. Furthermore, it also underscores the pivotal role of N in bolstering plant growth and biomass accumulation, offering valuable insights into optimizing nitrogen management strategies to enhance vegetation productivity.
Nonstructural carbohydrates (NSC) regulate physiological stress and serve as substrates for plant growth and metabolic processes (Ouyang et al., 2016; Liu et al, 2018a; Liu et al., 2018b). The NSC flow and distribution among plant organs reflect the delicate balance between carbon uptake and consumption (Aubrey and Teskey, 2018). Research has demonstrated that adding N and P can influence the NSC and its distribution in entire plant organs (Xiao et al., 2017; Li et al., 2021). During NSC synthesis, leaves serve as the primary organs. Following synthesis, NSC initially fulfills the requirements of the leaf before being gradually transported downwards and ultimately stored in the plant root. N and P addition enhances leaf capacity to produce NSC, resulting in a typical NSC content order of leaves > roots > stems (Li et al., 2016; Mo et al., 2020). Soluble sugars, acting as signaling molecules, enable cells to adapt to environmental fluctuations and participate in cellular osmoregulation. The soluble sugar content reproduces the plant's capability to adapt to its environment to some extent (Ouyang et al., 2016; Mo et al., 2020. However, there are discrepancies regarding the influence of N and P application on plant NSC accumulation and distribution. For instance, (Liu et al., 2021) reported decreased soluble sugar content with increasing N addition. (Xie et al., 2018) observed variable leaf-soluble sugar content responses to N addition across different plant species. Similarly, the impact of P addition on plant nutrient partitioning exhibits varying results; Li et al. (2016) observed an increase in leaf NSC content, where a decrease was reported by Wu et al. (2022a). Consequently, the precise impact of N and P on NSC accumulation and distribution remains inconclusive, necessitating further investigation (Han et al., 2005; Wu et al., 2022a).
Earlier studies have predominantly focused on assessing the effects of N and P supplementation on plant leaf nutrient content, photosynthetic activity, and hydraulic characteristics, often overlooking the significance of other vital plant parts (Han et al., 2005; Wu et al., 2022a,b). However, it is crucial to recognize that responses observed at the leaf level might not accurately mirror the reactions occurring across the entire plant on a broader spatial and sequential scale (Wu et al., 2022a,b; Li et al., 2023). This limitation underscores the necessity for investigations that encompass a more comprehensive perspective. The present study was conducted within a subtropical broad-leaved evergreen forest to address knowledge gap. Over nine consecutive years, the study evaluated the responses of naturally regenerated juvenile trees, particularly for the dominant Castanopsis sclerophylla species within the stand. Increased N deposition was hypothesized to significantly boost plant biomass accumulation through heightened leaf N content and enhanced photosynthetic pigment synthesis, leading to increase above-ground growth. Moreover, studied mentioned limited understanding of the combined effects of nitrogen (N), phosphorus (P), and water on plant biomass allocation and growth, as most studies have focused on individual nutrients or environmental factors (Wu et al., 2021).Unclear mechanisms of how N and P additions affect the distribution of non-structural carbohydrates (NSC) across plant organs, particularly how soluble sugar content varies among species. A need for long-term studies that encompass the whole plant response to nutrient additions, as previous research often focused only on leaf-level responses, overlooking the broader plant dynamics (Zhou et al., 2021). We hypothesize that N and P addition will significantly enhance plant biomass accumulation by increasing leaf N content and photosynthetic pigment synthesis, particularly in dominant species like Castanopsis sclerophylla. P addition is expected to alter the distribution of non-structural carbohydrates (NSC) across plant organs, leading to variations in NSC content between leaves, roots, and stems. Additionally, the effects of N and P on nutrient allocation and NSC distribution will vary among species, reflecting changes in soluble sugar content and nutrient cycling dynamics in subtropical ecosystems. These hypotheses aimed to investigated long-term impacts on forest ecosystem dynamics by studying the responses of dominant tree species, providing insights into nutrient cycling and predicting future vegetation evolution in the subtropical forest. Ultimately, these findings are helpful to serve as a foundational reference for predicting the trajectory of vegetation success and anticipating the potential alterations in forest characteristics within future habitats.
2 Materials and methods
2.1 Study area
The experimental site was situated within the Rending Mountain Forest Farm, Shitai County, Chizhou City, Anhui Province, China (117° 26′ 24″ N, 30° 15′ 37″ W), at an elevation of 120 m. This region experiences a north subtropical monsoon climate, characterized by high temperatures during summer, an annual mean temperature of 16.1 °C, peaked at 38.8 °C recorded in recent years. The County receives mean yearly precipitation of 1,626 mm, accompanied by 1,704.4 annual hours of sunshine and a frost-free period lasting 234 days, on average. The soil was predominantly sandy-yellow–red loam. The vegetation comprised a broad-leaved evergreen forest, with dominant species including C. sclerophylla, Quercus acutissima, and Castanopsis eyrei. The area primarily consisted of secondary forests of C. sclerophylla covering approximately 500 acres, largely regenerated naturally following logging activities in the 1960 s and subsequent forest management practices over the past five decades.
2.2 Plot design
In August 2011, we established 12 plots (15 m × 15 m) based on four treatments and three replicates within a 50-year-old C. sclerophylla secondary forest. To prevent nutrient infiltration interference from runoff between plots, buffer zone of about 10 m was maintained between them, delineated by signs and pull ropes. A randomized block design was used for the experiment in 12 plots, applying four treatment under different levels subjected 3 plots designated for each treatment (Zheng et al., 2016). The detail of different nutrient addition included N + P, N, P, and CK is given in Table 1. N and P were administered via ammonium nitrate (NH4NO3), and calcium was traced in superphosphate [Ca(H2PO4)2], respectively. We dissolved these fertilizers in 20 L of water and sprayed throughout the forest using artificial sprayer, while CK plots received equal water spray only.
Treatments
Single additions (kg)
Equivalent inputs (kg ha−1 year−1)
Mean DBH (cm)
NH4NO3
Ca(H2PO4)2
N
P
(N + P) addition
1.61
5.184
100
50
21.43 ± 0.47
N addition
1.61
0
100
0
21.26 ± 0.46
P addition
0
5.184
0
50
20.98 ± 0.72
CK (control)
0
0
0
0
18.93 ± 0.50
2.3 Plant growth measurement, sample collection, and processing
In August 2020, ten seedling of C. sclerophylla of one year age that are naturally regenerated show comparable vigor were carefully selected within natural secondary forest. Before harvesting, precise measurements of height (m) and diameter (D) were taken and recorded for these young seedlings using ordinary tap and caliper (Li et al., 2024a), focusing on the area near the soil's surface. These selected samples were carefully dried under controlled temperature of 70 °C until a constant weight was achieved, and their biomas (dry weight) were precisely determined. Then, samples were accurately sieved, to enable the determination of their C, N, and P contents using standard methods (Li et al., 2023). Specifically, the Walley-Black's wet digestion method was employed for C analysis, the Kjeldahl method was utilized for N determination, and the molybdenum-blue colorimetric method was used for P quantification (Li et al., 2024b). An assessment of soluble sugars and starch contents was also conducted Mitchell et al. (2013), using ethanol to extract soluble and subsequently quantified through the anthrone colorimetric approach (Kejla et al., 2023). The nonstructural carbohydrate (NSC) content was calculated as the accumulative total of each sample's soluble sugars and starch contents (Zhang et al., 2024).
2.4 Measurement of gas exchange parameters
In August 2020 and January 2021, selection of ten seedlings of C. sclerophylla that exhibited similar growth characteristics. This selection was made under each treatment condition (CK, N, P, and N + P) during sunny weather, particularly in the morning from 9:00 to 11:00 am. To determine key physiological traits such as the stomatal conductance (mol m-2 s-1), leaf net photosynthetic rate (μmol m-2 s-1), and transpiration rate, 3–5 mature healthy, leaves faced to sunlight were chosen from selected seedlings. We sued precise portable photosynthesis meters (model Li-6400 XT, LI-COR) for measuring these factors (Wu et al., 2022a; Wu et al., 2022b). Before taking the measurements, the experimental conditions were predetermined. Specifically, the light intensity was fixed at 1500 μmol mol m−2 s−1,the leaf chamber temperature was maintained at 28 °C, and the CO2 concentration was kept constant at 400μmol mol-1. All measurements were conducted on the same day to ensure consistency and accuracy. Before recording any data, the collected plant leaves were stabilized within the leaf chamber for 5–10 min, ensuring that each gas exchange parameter reached a stable state.
2.5 Statistical analysis
In this comprehensive study, we applied multi-way ANOVA to evaluate the effects of different factors. Additionally, the t-test identifies significant differences in these parameters between N and P alteration treatments compared to CK, with a significance standard level set at α = 0.05, was applied. To gain further insight into plant NSC characteristics and identify the key factors that shape them, redundancy analysis (RDA) and Monte Carlo tests were performed using Canoco 5 software. Statistical software like SPSS 20.0 were used for data analysis to visualize variations among the treatments, while GraphPad Prism 8 software and Microsoft Excel 2010 were used for graphical representations. In addition, we tested the data applying Pearson's correlation coefficient to show statistical relations among elemental contents of C, N, and P, and their ratios (C:N, C:P, N:P), starch, soluble sugars, soluble sugars, NSC contents, and starch ratio.
3 Results
3.1 Effects of N and P on nutrient stoichiometry in C. sclerophylla seedlings
In the analysis, multifactorial ANOVA showed that elemental contents (C, N, and P) significantly (p < 0.01) affected seedling organs and their associated seasonal relationship (Fig. 1, Table 2). During wet season, root absorbed N showed a significant higher intake under CK (p < 0.05) than combined N + P addition. However, stem N content was notably higher under N treatment compared to CK, P, and combined N + P treatments. While, N content in the branch content was significant (p < 0.05) under the P addition and N addition. During the dry season, the foliar P content under combined N + P treatment was found to be notably higher (21.66 %) and 34.34 % more in the CK. Comparatively, the wet season, the N significantly (p < 0.05) under the same treatment of N + P in the dry season. However, the branch's P content under N + P and P addition notably higher than in the dry season (p < 0.05). In contrast, N addition contributed to the P content of branches, which was significantly higher than combined N + P, P applications, and CK in the wet season, ranging from 58.02 % to 128.27 % (p < 0.05).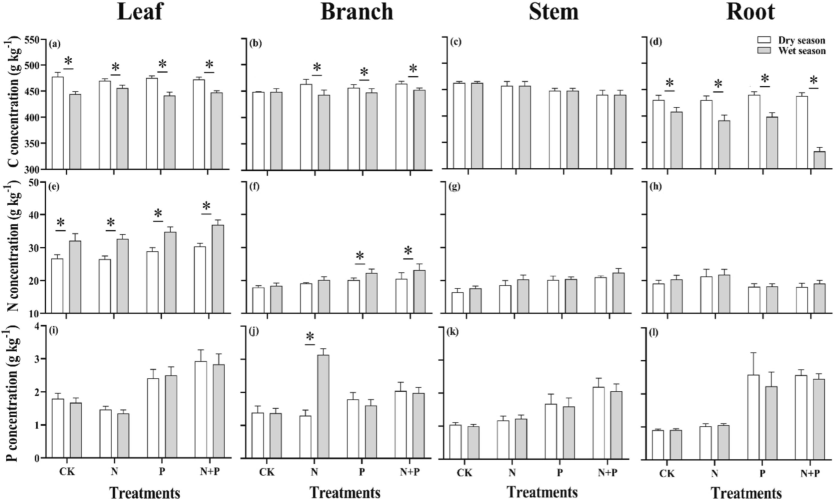
C, N, and P content in different organs of Castanopsis sclerophylla seedlings to N and P addition during the wet and dry seasonl; Leaf C,N,P (a,e,i), branch C,N,P (b,f,j), stem C,N,P (c,g,k), Root C,N,P (d,h,i), Asterisks indicate significant differences between the same treatments across seasons.
Parameters
T
S
O
T × S
T × O
S × O
T × S × O
C
0.642
0.000
0.000
0.708
0.112
0.000
0.686
N
0.001
0.000
0.000
0.941
0.002
0.000
0.998
P
0.000
0.653
0.000
0.966
0.008
0.970
1.000
C:N ratio
0.000
0.000
0.000
0.972
0.000
0.369
0.994
C:P ratio
0.000
0.493
0.000
0.859
0.002
0.846
0.996
N:P ratio
0.000
0.018
0.015
0.980
0.000
0.189
0.916
Soluble sugars
0.023
0.185
0.000
0.603
0.001
0.716
0.905
Starch
0.002
0.003
0.000
0.963
0.000
0.414
0.971
NSC
0.001
0.003
0.000
0.902
0.000
0.682
0.974
Soluble sugars: Starch
0.886
0.000
0.000
0.584
0.010
0.000
0.693
Different treatments, seasons, organs, and their exchanges significantly (p < 0.05) affected the elemental ratio (C:N, C:P, and N:P) of C. sclerophylla seedlings (Table 2). Applying N, P, and N + P, considerably higher (p < 0.05) CK was found in the wet season (Fig. 2). The C:P ratio of roots in the dry and wet seasons had the same trend; specifically, the N-added and CK were more significant (p < 0.05) than other treatments. The leaf N:P ratio under the N addition in the dry season was significant compared to CK and P addition, and the CK and P addition showed a higher ratio under N + P addition. In the wet season, the leaf N:P ratio under the N addition was significantly higher than the CK (p < 0.05). The stem N:P ratio was substantially higher in the dry and wet seasons under the N addition (p < 0.05). Under the N addition and CK, the root N:P ratio was significantly higher (p < 0.05) than the N + P and P additions in the dry and wet seasons.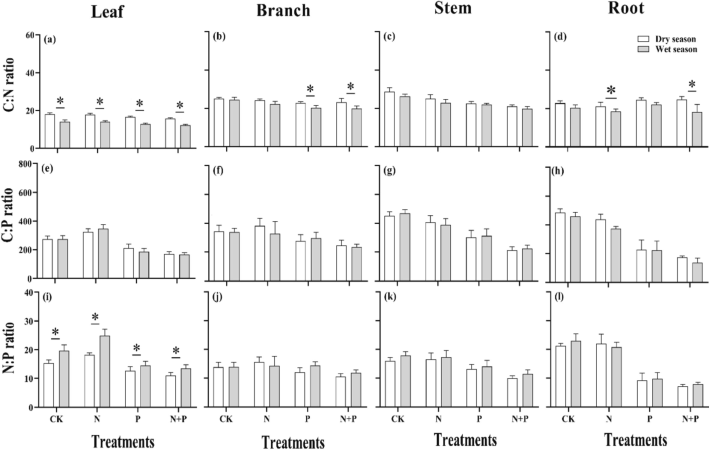
C:N, C:P, and N:P ratios in different organs of Castanopsis sclerophylla seedlings to N and P addition during the wet and dry season; Leaf C:N, C:P,N:P ratio (a, e,i), branch C:N, C:P,N:P ratio (b.f.j), stem C:N, C:P,N:P ratio (e,g,k), root C:N, C:P,N:P ratio (d,h,i), Asterisks indicate significant differences between same treatments across seasons.
3.2 Effects of N and P additions on NSC characteristics in C. sclerophylla seedlings
In the analysis, C. sclerophylla organ content and treatment-organ interaction showed significant (p < 0.01) affect on plant soluble sugars, starch, and NSC substances (Table 2). The seedlings branches soluble sugar content under N + P addition in the wet season was significantly higher (p < 0.05) than that of N, P, and control (CK) treatments in the dry season (Fig. 3). Overall, in both dry and wet seasons, root soluble sugar content showed a significant increase (p < 0.05) in the P-added and CK treatments than in the N (23.49 %) and N + P additions (34.17 %). Leaf starch content under N, P-added addition was significantly higher (p < 0.05) compared to the only P-added addition and CK in the wet season. Root starch content was highest under the N + P addition in both seasons and was higher (p < 0.05) compared to the CK, ranging from 45.60 % to 58.70 %.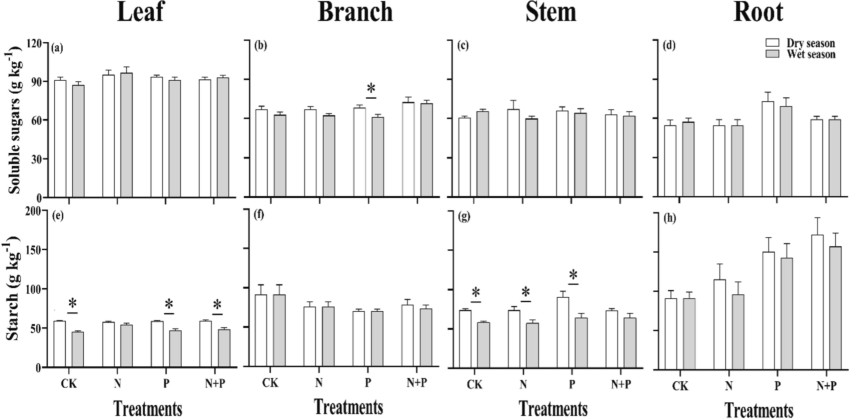
Effects of N, P addition on soluble sugars and starch content in different organ of Castanopsis sclerophylla seedlings during the wet and dry season, Leaf soluble sugar, starch (a,e), branch soluble sugar, starch (b,f), stem soluble sugar, starch (e,g), root soluble sugar, starch (d,h), Asterisks indicate significant differences between same treatments across seasons.
Under the N + P and N additions, The leaf NSC content of was significantly higher in the wet season and found lower in the dry season under the P-added addition and CK (Fig. 4). The NSC content was significantly higher (p < 0.05) than the N, P, and N + P additions in the wet season in the. Soluble sugars to starch ratio in foliar under the N + P, P addition, and CK was considerably higher (p < 0.05) than in the dry season. The starch content of branches under the N addition was significantly low (p < 0.05) in different seasons, and the stem soluble sugars to starch ratio was significantly higher (p < 0.05) in the dry season. Significantly, the stem soluble sugars to the starch ratio in the dry season were higher than in the wet season (p < 0.05).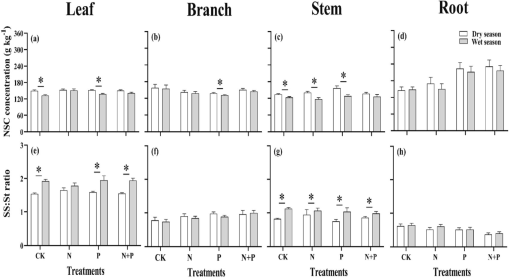
Effects of N, P addition on NSC concentrations and soluble sugars:starch ratio in different organ of Castanopsis sclerophylla seedlings during the dry and wet season, leaf NSC, SS:st ratio (a,e), branch NSC, SS:st ratio (b,f), stem NSC, SS:st ratio (e,g), root NSC, SS:st ratio (d,h), Asterisks indicate significant differences between same treatments across seasons.
3.3 Effects of N and P additions on photosynthetic parameters in C. sclerophylla seedlings
Transpiration rate and stomatal conductance of the C. sclerophylla seedlings in different treatments varied significantly (p < 0.05) in seasons (Table 3). The photosynthesis rate under the N, P-added addition, N-added addition, and CK were significant in the dry season (p < 0.05). At the same time, it was more important than the P-added addition (p < 0.05) in the wet season. Overall, seasonal differences affect the photosynthesis rate, which increased in the wet season compared to the dry season (p < 0.05) (Fig. 5).
Parameters
T
S
T × S
Photosynthetic rate
0.182
0.048
0.385
Transpiration rate
0.000
0.000
0.747
Stomatal conductance
0.012
0.000
0.789
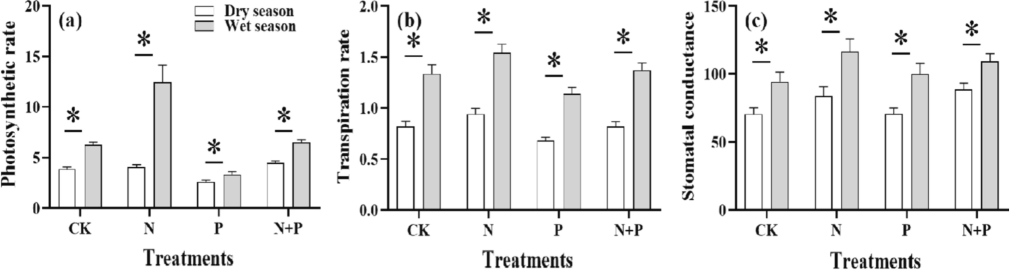
Effects of N, P addition on photosynthetic rate, transpiration rate, and stomatal conductance in different organs of Castanopsis sclerophylla seedlings during the wet and dry season, photsysthesis rate (a), transpiration rate (b), stomatal conductance (c), Asterisks indicate significant differences between the same treatments across seasons.
The transpiration rate to the photosynthesis rate in wet and dry seasons maintained similar trends. There was no significant difference (p > 0.05) concerning stomatal conductance under different additions in the wet season. However, the seasonal difference in stomatal conductance under all additions was higher in the wet season (p < 0.05).
3.4 Correlation analysis between C, N, and P stoichiometry and NSC characteristics of C. sclerophylla seedlings
The leaf C in C. sclerophylla seedlings exhibited a statistically significant positive correlation with soluble sugars and to the ratio of soluble sugars to starch (p < 0.05). Conversely, there is a significant negative correlation with starch content (p < 0.05) (Fig. 6). Similarly, leaf N content displayed a significant positive and negative correlation. Leaf P content correlated positively with soluble sugar content (p < 0.05). The leaf C:P ratio demonstrated significant negative association with soluble sugar content and the N:P ratio. At the same time, there was a negative correlation with NSC content (p < 0.05).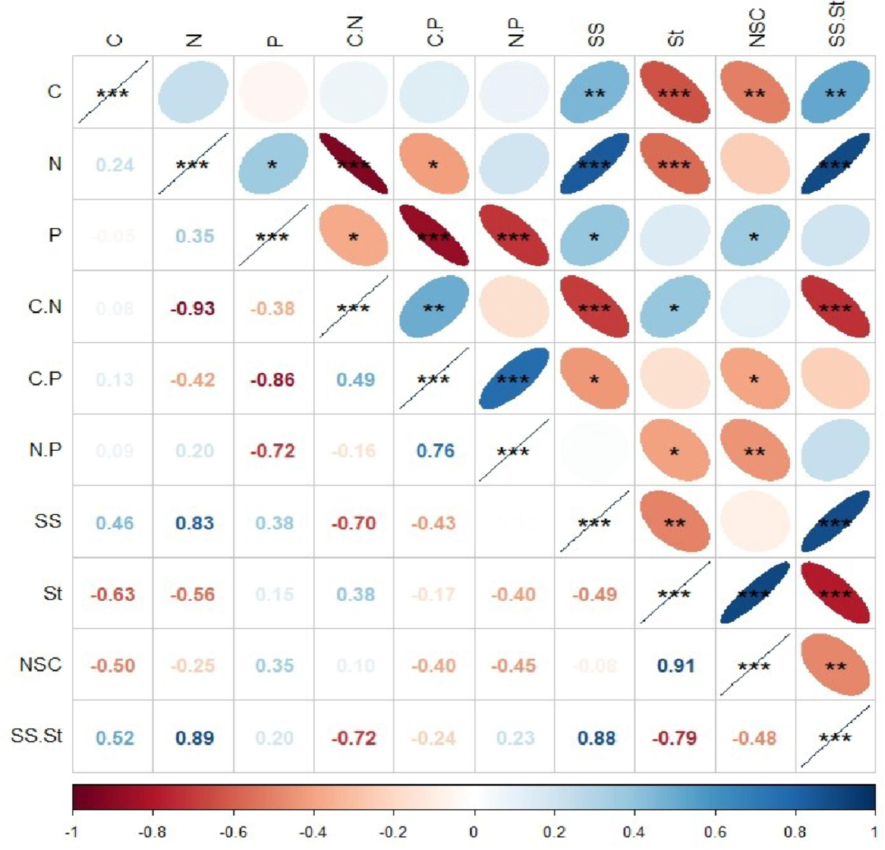
Correlation between C, N, P and NSC characteristics of leaf Castanopsis sclerophylla seedlings in different treatments. Note: C, carbon; N, nitrogen; P, phosphorus; SS, soluble sugars; St, starch; NSC, nonstructural carbohydrates. ***, p < 0.001; **, p < 0.01; *, p < 0.05.
Statistically, this study determined the significance of plant C, N, and P trait factors on NSC traits under environmental factors. The magnitude of importance followed the order of C > P > C:P > C:N > N:P > N, with C and P content exerting a significant influence on NSC traits (p < 0.05). During the wet season, the eigenvalues of sorting axes 1 and 2 were 0.8118 and 0.1129, respectively. Similarly, as shown in Fig. 7, Monte Carlo showed the essential role of plant carbon, nitrogen, and phosphorus trait factors in NSC size traits. Results revealed that the order of importance was C > N > N:P > C:P > C:N > P, with C and N content exerting a highly significant influence on NSC traits (p < 0.05).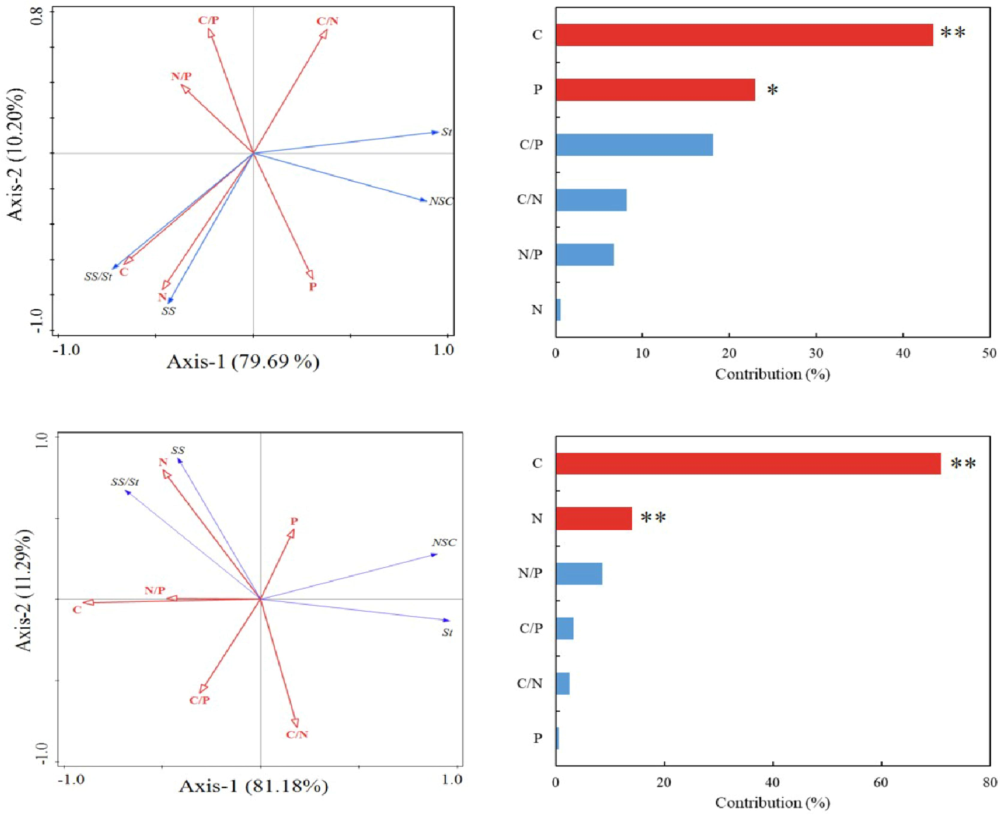
C, N, P, and NSC characteristics of leaf Castanopsis sclerophylla factor redundancy analysis. Note: C, carbon; N, nitrogen; P, phosphorus; SS, soluble sugars; St, starch; NSC, nonstructural carbohydrates. Indicators that could not further improve the goodness-of-fit have been excluded. **, p < 0.01.
4 Discussion
4.1 N and P effects on photosynthesis and plant growth
The forest regeneration growth stage of trees represents the most delicate and responsive phase in the lifecycle of forest trees. The effect of applying N and P on trees, particularly about tree age, remains focus of resecent research. Nevertheless, some studies have suggested a stimulatory effect, indicating positive influence on tree growth and development (Zheng et al., 2016; IPCC et al., 2018; Feng and Zhu, 2021). While other studies have shown an inhibitory effect, the response of young trees after nutrient addition is related to the tree species and the availability of light, water, and other site resources (Guan and Wen, 2011; Liu et al., 2019), promote chlorophyll synthesis and enhance leaf. However, in this study, photosynthesis did not respond to nutrient addition. This may be due to the high degree of canopy closure (> 0.8) in the C. sclerophylla stand, resulting in the understory young trees being limited by light availability (Guan and Wen, 2011; Liu et al., 2018a). Our results showed that addition N and P had no significant effect on the growth of one-year-old C. sclerophylla seedlings. In other words, the seedlings did not have enough light energy to drive CO2 assimilation in the understory forest. However, an increase in NSC content showed slight effect of young trees to the different nutrients absorbed, which may stimulate their growth potential in the future under better stand conditions (Zak et al., 2000).
4.2 Effects of N and P on stoichiometric traits of seedlings
Leaf N content significantly influences the photosynthetic rate of plants. N deficiency can increase inhibitors, delaying the photosynthetic process and efficiency (Wu et al., 2022a; Wu et al., 2022b). Decreased N content can reduce chlorophyll and enzyme activity, decrease photosynthetic rate, and cause metabolic disorders (Warren et al., 2000), P is the second most crucial element affecting photosynthesis and plant growth, plays a pivotal role in the structure of photosynthetic enzymes. Changes in leaf P content can notably impact the photosynthetic capacity of plants (Thomas et al., 2006). Elser's (2007) study found a negative relationship between plant leaf N and P content with global mean temperature. The findings indicated that, regardless of the wet or dry season, the mean N content in the CK group of C. sclerophylla seedlings exceeded the national scale value of 20.24 mg g−1 and the global vegetation level of 20.10 mg g−1. Similarly, the mean leaf P content was higher than the standard global value but lower than that reported by 42 plant species in southeastern China (2.24 mg g−1), with a comparison to the leaf P content of the north–south sample zone of eastern China being 1.77 and 1.28 mg g−1, respectively (Wu et al., 2012; Wang and Zheng, 2021; Wu et al., 2022a).
Plant leaf N:P ratios are critical for understanding nutrient limitations, vegetation composition, and ecosystem functioning in changing environmental conditions. Güsewell (2004) emphasized that Foliar ratio of N:P exceeding 16 often signifies P-limited plant growth. Our study revealed that C. sclerophylla seedlings in the CK exhibited leaf N:P ratios well above 16 during the wet season, whereas the ratios were below 16 in the dry season. Notably, in the wet season, leaf N content saw significantly higher leaf N content tha, leaf N content was significantly higher than in the dry season under CK conditions. In contrast, leaf P content remained relatively stable between the two seasons. This suggests that variations in leaf N:P ratios may be influenced by differences in leaf N content, indicating a more significant effect of N addition on C. sclerophylla seedlings during the wet season, confirmed in the previous study (Wu et al., 2022a). This observation aligns with the seedlings' growth patterns. However, adding P and N + P significantly reduced leaf N:P ratios in the wet and dry seasons compared to those under N addition. This alleviated the nutrient imbalance caused by prolonged N deposition. Subtropical forests are typically P-limited, and extended N deposition exacerbates P limitation in the soil (Wang et al., 2023). Plants may respond by increasing their uptake of external P to meet their growth needs, thus mitigating the long-term P-limiting pressure and alleviating nutrient imbalances (Zheng et al., 2016). This finding also implies that P inputs can be beneficial in mitigating the adverse effects of prolonged N sedimentation on plants (Wu et al., 2022a).
4.3 Effects of N and P on NSC content distribution in seedlings
The soluble sugar content within plants is intricately linked to their resilience against adverse environmental conditions (Wu et al., 2012). Storing C in harsh environments serves a survival function rather than supporting growth (Xing et al., 2023). Fluctuations in leaf NSC levels reflect the combined influence of soluble sugars and starch. The NSC content augmentation observed in C. sclerophylla leaves is primarily attributed to significant rises in soluble sugar and starch content. This NSC content increases and balances cellular osmotic pressure (Smith and Stitt, 2007), allowing the plant to adapt to environmental stresses. Stored C will be utilized during a C deficit until reserves are depleted (Poorter and Kitajima, 2007). However, starch accumulation is attributed to immobility, whereas soluble sugars can be redirected towards various physiological and metabolic plant activities. Consequently, leaves and root systems exhibit the highest soluble sugar content (Chantuma et al., 2009). Photosynthesis, a highly active physiological process, is plants' primary starch source, explaining the lower starch content observed in C. sclerophylla leaves.
The NSC content within plant organs significantly impacts the plant's ability to respond to environmental stresses, indicating carbon supply, buffering capacity, plant growth, and adaptive strategies (Zhang et al., 2024). In wet-season C. sclerophylla seedlings, NSC levels were notably higher under N + P and N addition compared to P addition and CK. This observation aligns with previous findings from subtropical forest studies examining the effects of N and P supplementation (Blumstein et al., 2022; Zhang et al., 2024). As a direct product of photosynthesis, NSC exhibits a strong positive correlation with plant photosynthetic capacity. A heightened photosynthetic rate facilitates NSC accumulation. However, in the wet season, with phosphorus addition, leaf photosynthesis of C. sclerophylla seedlings may be unable to sustain the higher respiration and rapid growth demands. Consequently, leaf starch is converted into soluble sugars to support the plant's needs. This accounts for the decreased starch content under phosphorus addition during the wet season. Furthermore, N and N + P addition elevated the soluble sugar content in C. sclerophylla seedling leaves, potentially related to the plant's response to drought stress caused by Warming. Additionally, NSC content in the root system increased under N and N + P addition, potentially enhancing the seedlings' drought resistance in dryer environments.
5 Limitations
While this study provides valuable insights into the effects of N and P additions on nutrient stoichiometry, NSC content, and photosynthetic parameters in C. sclerophylla seedlings, several limitations this study considered. First, this research focused on a relatively short time frame, and the long-term effects of nutrient additions on seedling development remain unclear. Additionally, the experimental design did not account for potential interactions between nutrient availability and other environmental factors, such as soil moisture or microbial activity, which could influence nutrient uptake and metabolism. Lastly, the study was conducted in an open forest environment, and field conditions may present different responses due to varying environmental stresses.
6 Conclusions
The addition of N, P, and combined N + P significantly increased the N and P content in the stem, leaf, root, and branches of C. sclerophylla seedlings. The plants actively absorbed these nutrients and utilized them for growth and development. Moreover, the interaction between nutrient additions and seasonal variations produced a coupling effect, stimulating a strong plant response. The NSC content of seedlings' root system increased under N + P additions and N additions alone. Results further suggest that the foliar elements N, C, and P contents in regulating the soluble sugars and starch metabolism in C seedlings. The negative correlation between the ratio of C:P and NSC contents indicates that maintaining a balanced nutrient status is essential for optimal carbohydrate metabolism in these plants. These findings are critical for establishing a foundational reference for forecasting the path of vegetation development in the region under alterations in forest characteristics in the near future. Further this suggest that targeted nutrient management strategies, particularly the balanced addition of N and P, could enhance the growth and resilience of C. sclerophylla regrowth in response to environmental changes. Furthermore, understanding the role of nutrient stoichiometry in carbohydrate metabolism may aid in predicting how forest ecosystems will adapt to climate-driven shifts in nutrient availability and seasonal dynamics.
CRediT authorship contribution statement
Zhaocheng Wang: Writing – original draft, Resources, Project administration, Methodology, Formal analysis, Conceptualization. Xu Li: Software. Muhammad Arif: Writing – review & editing. Jaweria Shamshad: Writing – review & editing. Aimin Wu: Software. Wenyong Zhan: Project administration. Bilal Ahmad: Writing – review & editing. Nengzhi Tan: Visualization, Supervision, Funding acquisition, Conceptualization. Khalid Mashay Al-Anazi: Writing – review & editing. Mohammad Abul Farah: Writing – review & editing. Muhammmad Sadiq Khan: .
Funding
This research received no external funding.
Acknowledgments
This research was funded by Huzhou Natural Science Foundation Project (2022YZ19); Huzhou Vocational and Technical College High Level Talent Project (2022ZS09) and the Anhui Postdoctoral Fund Project (2022B582). The authors would like to extend their sincere appreciation to the Researchers Supporting Project number (RSP2024R154), King Saud University, Riyadh, Saudi Arabia.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Stored root carbohydrates can maintain root respiration for extended periods. New Phytol.. 2018;218:142-152.
- [CrossRef] [Google Scholar]
- Plantcarbohydrate storage: intra- and inter-specific trade-offs reveal a major lifehistory trait. New Phytol.. 2022;235:2211-2222.
- [CrossRef] [Google Scholar]
- Carbohydrate storage in wood and bark of rubber trees submitted to different level of C demand induced by latex tapping. Tree Physiol.. 2009;29(8):1021-1031.
- [CrossRef] [Google Scholar]
- Carbohydrate dynamics and mortality in a pinon-juniper woodland underthree future precipitation scenarios. Plant Cell Environ.. 2015;38:729-739.
- [CrossRef] [Google Scholar]
- Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett.. 2007;10:1135-1142.
- [CrossRef] [Google Scholar]
- Global patterns and associated drivers of priming effect in response to nutrient addition. Soil Biol. Biochem.. 2021;153:1-11.
- [CrossRef] [Google Scholar]
- More nitrogen partition in structural proteins and decreased photosynthetic nitrogen-use efficiency of Pinus massoniana under in situ polluted stress. J. Plant Res.. 2011;124(6):663-673.
- [CrossRef] [Google Scholar]
- N: P ratios in terrestrial plants: variation and functional significance. New Phytol.. 2004;164(2):243-266.
- [CrossRef] [Google Scholar]
- Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol.. 2005;168(2):377-385.
- [CrossRef] [Google Scholar]
- IPCC. Summary for policymakers//Masson-Delmotte, V., Zhai, P.M., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani A., Moufouma-Okia, W., Péan C., Pidcock, R., Connors, S., Matthews, J.B.R., Chen, Y., Zhou, X., Gomis, M.I., Lonnoy, E., Maycock, T., Tignor, M., Waterfield, T., eds. Global Warming of 1.5℃. An IPCC Special Report on the Impacts of Global Warming of 1.5℃ Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty. Geneva, Switzerland: World Meteorological Organization, 2018.
- Anthrone method combined with adsorption of interferents as a new approach towards reliable quantification of total carbohydrate content in pyrolysis bio-oils. J. Anal. Appl. Pyrol.. 2023;173:106066
- [Google Scholar]
- Carbon sequestration and storage capacity of Chinese fir at different stand ages. Sci. Total Environ.. 2023;904:166962
- [CrossRef] [Google Scholar]
- Effects of nitrogen and phosphorus supply on growth and physiological traits of two Larix species. Environ. Exp. Bot.. 2016;130:206-215.
- [CrossRef] [Google Scholar]
- Mixed plantations do not necessarily provide higher ecosystem multifunctionality than monoculture plantations. Sci. Total Environ.. 2024;914:170156
- [CrossRef] [Google Scholar]
- Increasing stand age increases N deficiency but alleviates relative P limitations in Castanopsis hystrix plantations in Southern China. Land Degrad. Dev.. 2024;35(6):2173-2183.
- [CrossRef] [Google Scholar]
- Changes in carbon and nitrogen metabolism during seawater-induced mortality of Picea sitchensis trees. Tree Physiol.. 2021;41(12):2326-2340.
- [CrossRef] [Google Scholar]
- Responses of foliar carbohydrates and nutrient status of two distinctive cypress species to shading and nitrogen addition. Global Ecol. Conserv.. 2018;16:e00452.
- [Google Scholar]
- Nonstructural carbohydrates regulated by season and species in the subtropical monsoon broad-leaved evergreen forest of Yunnan Province. China. Scientific Report. 2018;8(1):1083.
- [CrossRef] [Google Scholar]
- Photosynthesis, ecological stoichiometry, and nonstructural carbohydrate response to simulated nitrogen deposition and phosphorus addition in Chinese fir forests. Forests. 2019;10(12):1068.
- [CrossRef] [Google Scholar]
- Effects of an eight-year nitrogen and phosphorous addition on leaf photosynthesis and chemistry of mature castanopsis sclerophylla trees in subtropical China. Applied Ecology & Environmental Research. 2021;19(5)
- [CrossRef] [Google Scholar]
- Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytol.. 2013;197(3):862-872.
- [CrossRef] [Google Scholar]
- Leaf nonstructural carbohydrate concentrations of understory woody species regulated by soil phosphorus availability in a tropical forest. Ecol. Evol.. 2020;10(15):8429-8438.
- [CrossRef] [Google Scholar]
- Response of foliar mineral nutrients to long-term nitrogen and phosphorus addition in a tropical forest. Funct. Ecol.. 2021;35:2329-2341.
- [CrossRef] [Google Scholar]
- Drought survival of tropical tree seedlings enhanced by nonstructural carbohydrate levels. Nat. Clim. Change. 2014;4(8):710-714.
- [CrossRef] [Google Scholar]
- Improvements in ecosystem services from investments in natural capital. Science. 2016;352:1455-1459.
- [CrossRef] [Google Scholar]
- Carbohydrate storage and light requirements of tropical moist and dry forest tree species. Ecology. 2007;88(4):1000-1011.
- [CrossRef] [Google Scholar]
- Coordination of carbon supply and plant growth. Plant Cell Environ.. 2007;30(9):1126-1149.
- [CrossRef] [Google Scholar]
- Leaf inorganic phosphorus as a potential indicator of phosphorus status, photosynthesis and growth of Eucalyptus grandis seedlings. For. Ecol. Manage.. 2006;223(1–3):267-274.
- [CrossRef] [Google Scholar]
- Nitrogen- or phosphorus-limited growth in herbaceous, wet vegetation: relations with atmospheric inputs and management regimes. Trends Ecol. Evol.. 1996;11:494-497.
- [CrossRef] [Google Scholar]
- Long-term nitrogen deposition inhibits soil priming effects by enhancing phosphorus limitation in a subtropical forest. Glob. Change. Biol.. 2023;29(14):4081-4093.
- [Google Scholar]
- Impact of vegetation succession on leaf-litter-soil C:N: P stoichiometry and their intrinsic relationship in the Ziwuling Area of China's Loess Plateau. J. For. Res.. 2021;32(2):697-711.
- [CrossRef] [Google Scholar]
- Is photosynthesis related to concentrations of nitrogen and Rubisco in leaves of Australian native plants? Aust. J. Plant Physiol.. 2000;27:407-416.
- [CrossRef] [Google Scholar]
- Direct and indirect effects of long-term field warming methods on the physical environment and biological responses in a subtropical forest. Front. For. Global Change. 2022;5:877025
- [CrossRef] [Google Scholar]
- Impacts of competition and nitrogen addition on plant stoichiometry and non-structural carbohydrates in two larch species. J. For. Res.. 2021;32:2087-2098.
- [Google Scholar]
- Physiological traits and response strategies of four subtropical tree species exposed to drought. Environ. Exp. Bot.. 2022;203:105046
- [CrossRef] [Google Scholar]
- Leaf nitrogen and phosphorus stoichiometry across forty-two woody species in Southeast China. Biochem. Syst. Ecol.. 2012;44:255-263.
- [CrossRef] [Google Scholar]
- Nitrogen addition has a stronger effect on stoichiometries of nonstructural carbohydrates, nitrogen and phosphorus in Bothriochloa ischaemum than elevated CO2. Plant Growth Regul.. 2017;83(2):325-334.
- [CrossRef] [Google Scholar]
- Leaf non-structural carbohydrate allocation and C: N: P stoichiometry in response to light acclimation in seedlings of two subtropical shade-tolerant tree species. Plant Physiology and Biochemistry. 2018;124:146-154.
- [CrossRef] [Google Scholar]
- Effects of long-term nitrogen addition on fine root dynamics in a temperate natural secondary forest. Scand. J. For. Res.. 2023;38(3):154-165.
- [CrossRef] [Google Scholar]
- Atmospheric CO2, soil-N availability, and allocation of biomass and nitrogen by Populus tremuloides. Ecol. Appl.. 2000;10(1):34-46.
- [CrossRef] [Google Scholar]
- Contrasting coordination of nonstructural carbohydrates withleaf and root economic strategies of alpine coniferous forests. New Phytol. 2024
- [CrossRef] [Google Scholar]
- Effects of phosphorus addition with and without nitrogen addition on biological nitrogen fixation in tropical legume and non-legume tree plantation. Biogeochemistry. 2016;131:65-76.
- [CrossRef] [Google Scholar]
- Thinning promotes the nitrogen and phosphorous cycling in forest soils. Agric. For. Meteorol.. 2021;311:108665
- [Google Scholar]
Appendix A
Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jksus.2024.103517.
Appendix A
Supplementary data
The following are the Supplementary data to this article:




