

Translate this page into:
Ameliorative effect of melatonin on different tomato genotypes to induce heat stress tolerance by modulating growth and physiological attributes
⁎Corresponding authors. rashad.mukhtar@uos.edu.pk (Rashad Mukhtar Balal), parvaizbot@yahoo.com (Parvaiz Ahmad)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Climate change; the most concerning issue throughout the globe, is now considered a major cause of heat stress which is deteriorating agricultural crops. A pot experiment was performed to examine the ameliorative role of melatonin foliar spray on growth attributes, physiological attributes and yield and quality attributes of heat-tolerant and heat-sensitive genotypes of tomatoes under heat stress mediated by field environment. The results demonstrated that all the growth parameters of all tomato genotypes such as plant fresh biomass, plant dry biomass, stem girth, leaf area, the number of viable seeds, protein contents as well as physiological attributes including photosynthetic rate, stomatal contents, transpiration rate, chlorophyll contents, water use efficiency (WUE) and synthesis of osmoprotectants including proline and glycine betain (GB) were increased significantly upon foliar application of melatonin @ 25 µM under heat stress except electrolyte leakage (EL), leaf temperature and hydrogen peroxide contents (H2O2) which were decreased in all cultivars of tomato either in heat tolerant or in heat sensitive; as compared to their respective control which remained untreated. Similarly, acivity of enzymatic and non-enzymatic antioxidants including nitric oxide synthase (NOS), glutathione reductase (GR), nitrate reductase (NR) and glutathione S transferase (GST) were also improved upon melatonin foliar application under heat stress. However, the maximum improvement in all measured attributes was observed in all types of tomato genotypes grown under both control and 25 µM melatonine foliar spray treatment. All these findings proved that melatonin spray is capable to address the deteriorative impact of high temperature on tomato growth.
Keywords
Heat tolerance
Heat sensitivity
High temperature
Plant biomass
Photosynthesis rate
Transpiration rate

1 Introduction
A change in temperature; even more than 1 ℃, is able to produce heat stress. In specific climatic areas or zones, heat stress is considered as a high degree of temperature. It occurs at day or night and depends upon the climatic conditions, that raise the temperature to its optimum level. Heat-tolerant plants are those plants that can produce an economic yield of the crop under extreme temperatures. The rise in greenhouse gasses may gradually increase the global temperature (Ahammed et al., 2021). Due to the temperature rise, the growing season of plants may be altered, along with the disturbance in the geographical distribution. This rise in temperature to its threshold level allows it to damage the early crop maturity at the start of the season as well (Fahad et al., 2019). High-temperature stress may harm the microtubule organization and mitotic cells aster formation, phragmoplast microtubules and splitting of spindles may be damaged (Bellinger, 2020). Heat stress injuries cause the production of reactive oxygen species (ROS), toxic compounds, starvation, and growth inhibition in a plant cell (Hemantaranjan et al., 2018).
Heat stress may cause the actual changes in plants at the cellular level. These changes in the synthesis of proteins are related to stress which may change the gene expression level (Raja et al., 2020). During heat stress, some plants adapted heat shock proteins to cope with heat stress conditions. Like chaperone heat shock proteins (HSP), The molecular mass of HSPs ranges from 10 to 200 KD (kilo Daltons). HSP works in signal transduction when the condition is extreme to heat stress (Ul Haq et al., 2019). However, all plants with the same genotype or within the species are not capable of coping with heat stress conditions. There are so many variations in the plant genome, within or between the plant species which make plant capable to stand against extreme heat stress environment by genetic difference (Ul Hassan et al., 2021). As plant changes their behavior with respect to the environment, it is not easy to find their upper threshold level of temperature stress (Nievola et al., 2017). However a temperature above 35 degrees, negatively affected seed germination, fruit ripening, vegetative growth, and fruits. The higher threshold level for other plant species may be upper and lower than the 35 °C. Heat stress is considered as the main limiting factor for production of crop plants. High temperature is a very sensitive element for crop yield (Argosubekti, 2020).During seed filling, high exposure to heat may reduce yield, and seed weight per kilogram, and also expedite the rate of senescence. The reason is that due to heat stress plants cope with it may lead to a reduction of the photosynthetic rate, divert the resources and reproduce within limited factors. When immediately exposed to heat stress (30–35 ℃), the plant may be affected adversely like flower dropping at the blooming stage (Ferguson et al., 2021).
This study draws attention to adaptations and plant responses to heat stress conditions. The plant tolerates extreme conditions for its genetic improvement by adopting different strategies and procedures at cellular and subcellular levels. Reduction in plant growth and development starts when the temperature of the environment exceeds its level is called a threshold temperature. Through field experts and control laboratory procedures, the temperature level has been changed for plant development and growth.
Melatonin is a very beneficial compound and provides a protective shield for the growth of plants under abiotic stress as well (Ahammed and Li, 2022). It has the ability to cure the plants that were damaged by heat stress or by drought conditions (Shafi et al., 2021). Heat stress injuries plant and the major part of the plant is affected by rupturing of the plasma membrane, production of ROS species, low rate of photosynthesis, dehydration in plants and premature falling of seeds and flowers before blooming (Imran et al., 2021). In different plant species, the function of melatonin is also different as it provides a strong defense system by improving osmoprotectant including proline and Glycine betaine which play its major role in plant defense system. It also works as an electron receptor and reduces the oxidative stress (ROS species) in plants leading to promote seed germination and development. Melatonin receptor is also found in terrestrial plants, where it regulates stomatal opening and closure (Tan et al., 2015; Jou et al., 2019). However, it is hypothesized that optimum level of melatonin spray can ameliorate the depressing effect of heat stress by improving physiological and gas exchange parameters, osmoprotectant production and enzymatic antioxidant activities in all genotypes of tomato plants. Therefore present study was performed to determine the foliar impact of melatonin on morphological, yield, gas exchange and quality related attributes including osmoprotectants in all genotypes of tomato under heat stress.
2 Materials and methods
2.1 Experimental design
A pot trial was performed under two factor factorial randomized complete block design (RCBD) to evaluate the heat stress-mitigation effect of melatonin spray in four tomato cultivars. Tomato seeds of four cultivars (two heat tolerantand two heat sensitive) were sown. Pots of all tomato cultivars were placed in field under sunshine in order to face heat stress by plant. Two treatments were applied [control (without foilarspray) and melatonin @ 25 µM)] to all genotypes of tomato and each was replicated five times. Heat tolerant genotypes were included as T60 F1 and Supercash F1 while heat sensitive genotypes were named as Nagina and Naqeeb.
The seeds of tested genotypes of tomato were grown in 12-inch plastic pots filled with peat moss for growth medium. Half-strength Hoagland and Arnon (1950) nutrient solution, was applied as a source of nutrients. Pots were placed in the field of research area. The variations in climate of Sargodha city remained 41/5 °C and rainfall of 115/5 mm Whereas ideal temperature for tomato growth varies between 22–28 °C. Melatonin was applied at two growth stages of tomato i. one week before the maturity stage. ii. One week before the harvesting stage and following parameters were studied.
2.2 Growth parameters measurements
Data regarding growth attributes was collected at the end of stress, 40 days after the sowing of tomato seeds. Leaf samples and plant samples of individual replications were uprooted and washed with distilled water and attached particles of growing media were removed from the roots. After that plants blotted with filter paper for the removal of water present on leaves and roots.
Plant fresh biomass, plant dry biomass, leaf area, stem girth, photosynthesis rate (Pn), stomatal conductance, transpiration rate (E), water use efficiency (Y/Et), leaf temperature, chlorophyll contents, electrolyte leakage (EL), hydrogen peroxide (H2O2), soluble protein contents and number of viable seeds were observed according to standard procedures. Leaf area of plant was measured with the help of Leaf area meter (Ll-3100; LI-COR Inc., USA).
2.3 Physiological measurements
From top fully expanded 2nd leaf of a young plant from each replication of treatments was selected to record the photosynthetic rate, transpiration rate, and stomatal conductance with the help of photosynthesis measuring-system CI-340 transportable infrared gas analyzer (Analytical Development Company, Hoddesdon, England). This information was recorded between 10.00 and 12.00 A.M. of the day with subsequent modifications: atmospheric pressure 99.9 kPa, molar flow of air, per unit area of leaf 403.3 mmol m−2 S-1, water vapor pressure into chamber ranged from 6.0 to 8.9 m bar, leaf temperature ranged from 28.4 − 32.4 °C, ambient CO2 concentration 352 mol mol−1, PAR at surface of leaves was highest upto 1711 mol m−2 S-1 and ambient temperature was ranged from 22.4-27.9 °C.
Chlorophyll content meter (CCM-300, ADC Bioscientific, UK) was used to estimate chlorophyll contents from fully matured leaf samples of tomato plants. Water use efficiency was measured by using the following equation.
Where, Y=Crop yield and Et = Evapotranspiration rate.
2.4 Electrolyte leakage
Electrolyte leakage (EL) measures cell damage. It was determined by the method of Shi et al. (2006). 0.25 g leaf sample was kept in 25 ml deionized water for 24 h in test tubes. After 24 h time period, EC1 was measured by using electrical conductivity meter. Then test tubes were placed in water bath for 1 h at 90 ℃. After that, EC2 was measured again. Thus EL% was the determined by following equation.. EC1 / EC2 x 100.
2.5 H2O2 quantification
Analysis of hydrogen peroxide (H2O2) quantification was made by a method described by MacNevin and Urone (1953) with some modifications (Brennan and Frenkel, 1977; Rivero et al., 2007).
2.6 Leaf temperature
Leaf temperature was estimated by an Infrared thermometer (AmiciKart® Digital Laser IR Infrared Thermometer-GM320) from 11 am to 12 noon during the stress period.
2.7 Protein contents
The soluble protein contents were estimated through Bradford (1976) method using spectrophotometer UV (PG instrument T60).
2.8 Number of viable seeds per fruit
To measure the number of viable seeds, three fruits per plant were randomly selected and their seeds were counted manually by gently extracting seeds from tomato fruit through manual process and their viability was tested in petri dishes using Whatman filter paper wetted with double distilled water. The seeds that ruptured their seed coat and showed growth of their cotyledon were counted and considered viable seeds.
2.9 Enzyme assays
Activity of enzymatic antioxidants including NOS, GR, NR and GST was assayed by using their respective detection kits (Solarbio Life science, Beijing, China) according to protocol. Centrifugation rate and molar extinction was different for all determined enzymes. These kits were used after digestion and collection of supernatant solution of 0.1 g leaf sample.
2.10 Experimental design and statistical analysis
The data was analyzed through statistical methods illustrated by Gomez and Gomez (1984). To find the difference of significance between treatment means at P<0.05 (n = 5) Tukey HSD test was used. Data were evaluated through Statistix 8.1 software.
3 Results
3.1 Impact of melatonin foliar treatment on morphological attributes of tomato genotypes
The results (Table 1) showed that under the foliar application of melatonin, plant fresh biomass was increased as 41.62 % and 40.98 % in heat tolerant (T60 F1 and Super cash F1) genotypes of tomato respectively showing values as 651.99 g and 650.56 g of tomato when compared to their respective control. Similarly, fresh biomass in heat sensitive (Nagina and Naqeeb) genotypes was recorded as 473.09 g and 472.01 g respectively showing 34.11 % and 31.20 % increase than their respective control. However, all genotypes either heat tolerant or heat sensitive were increased significantly (p ≤ 0.05) under melatonin spray. However, heat tolerant genotypes of tomato were more responsive than heat-sensitive cultivars of tomato.
Treatments
Tolerant Cultivars
Sensitive Cultivers
T60 F1
Super cash F1
Naqeeb
Nagina
Plant fresh biomass (g)
Control
460.38 ± 23.02c
461.47 ± 23.53c
359.76 ± 19.79d
352.74 ± 18.70e
25 µM
651.99 ± 32.60a
650.56 ± 33.18a
472.01 ± 25.96b
473.09 ± 25.07b
Plant dry biomass (g)
Control
126.82 ± 5.96d
134.47 ± 6.72c
74.25 ± 3.71 h
79.17 ± 4.35 g
25 µM
174.89 ± 8.22b
186.16 ± 9.31a
94.38 ± 4.72f
101.22 ± 5.57e
Leaf area (cm2)
Control
172.24 ± 7.75c
166.59 ± 7.33d
126.45 ± 6.32e
126.40 ± 6.45e
25 µM
250.86 ± 11.29a
237.12 ± 10.43b
171.62 ± 8.58c
172.62 ± 8.80c
Stem girth (cm)
Control
1.50 ± 0.08e
1.55 ± 0.09d
1.15 ± 0.06 g
1.32 ± 0.07f
25 µM
2.13 ± 0.12b
2.25 ± 0.12a
1.50 ± 0.08e
1.77 ± 0.10c
Protein contents (µg/g FW)
Control
48.84 ± 1.71e
46.96 ± 1.60f
41.32 ± 1.61 g
40.38 ± 1.49 h
25 µM
65.55 ± 2.29a
62.30 ± 2.12b
52.16 ± 2.03c
50.01 ± 1.85d
Number of viable seeds
Control
88.28 ± 3.97d
90.16 ± 4.15c
80.77 ± 3.39 g
77.95 ± 3.12 h
25 µM
93.90 ± 4.23b
95.64 ± 4.40a
83.78 ± 3.52e
81.45 ± 3.26f
Similarly, maximum increase (38.44 %) in plant dry biomass of Super cash (Table 1) under the foliar application of melatonin was recorded (186.16 g) as compared to without application of melatonin i.e. control (134.47 g). followed by T60 F1 presented the 37.90 % increase in dry biomass (174.89 g) as compared to control (126.82 g). Similarly, heat-sensitive genotypes of tomato expressed that greater plant dry biomass was observed in Nagina (101.22 g) as compared to control (79.17 g) showing 27.85 % increase in plant drybiomass. Whereas, Naqeeb showed 27.11 % improvement inplant dry biomass (94.38 g) as compared to control (74.25 g).
The results (Table 1) showed that under the foliar application of melatonin, 45.65 % increased leaf area was observed in T60 F1 (250.86 cm2) as compared to without application of melatonin i.e. control (172.24 cm2). Whereas Super cash F1 showed 42.33 % increase in leaf area (237.12 cm2) when compared with the control (166.59 cm2). In the case of heat-sensitive genotypes of tomato, the results revealed a greater leaf area in Nagina (36.56 %) and Naqeeb (35.72 %) (heat sensitive) cultivars than their respective control.
The results revealed that the maximum (45.16 %) increase in stem girth (Table 1) under the foliar application of melatonin was noted in Super cash F1 (2.25 cm) as compared to without application of melatonin i.e. control (1.55 cm). Whereas T60 F1 under foliar application of melatonin presented 42 % increase in stem girth (2.13 cm) as compared to the control (1.50 cm). Similar to heat-tolerant genotypes of tomato, foliar application of melatonin also increased the stem girth in heat-sensitive genotypes of tomato and results expressed 35.60 % increase in stem girth in Nagina (1.77 cm) as compared to control (1.32 cm). Whereas, Naqeeb under foliar application of melatonin showed 30.43 % impeovement in stem girth (1.50 cm) as compared to control (1.15 cm).
3.2 Impact of melatonin foliar treatment on quality attributes of tomato genotypes
The greater production of protein (Table 1) from heat-tolerant genotypes of tomato was observed in T60 F1 (65.55 µg g−1 FW) as compared to without application of melatonin i.e. control (48.84 µg g−1 FW) showing 34.21 % increase over control treatment. Whereas Super cash F1 under foliar application of melatonin showed the 32.66 % increase in protein contents (62.30 µg g−1 FW) as compared to control (46.96 µg g−1 FW). In the case of heat-sensitive genotypes of tomato, the 26.52 % increase inprotein contents were recorded in Naqeeb (52.16 µg g−1 FW) as compared to control (41.32 µg g−1 FW). While Nagina under foliar application of melatonin presented only 23.84 % improvement in protein contents (50.01 µg g−1 FW) as compared to its control (40.38 µg g−1 FW). The foliar treatment of melatonin significantly (p ≤ 0.05) increased the protein contents of all growing cultivars either heat tolerant or heat sensitive.
The foliar treatment of melatonin significantly (p ≤ 0.05) increased (Table 1) the number of viable seeds per fruit in all cultivrs of tomato (Table 1). The similar trend of results was observed in the case of the number of viable seeds per fruit of tomato. The maximum increase in (6.07 %) number of viable seeds per fruit from heat-tolerant genotypes of tomato was observed in Super cash F1 (95.64) as compared to without the application of melatonin (90.16). Whereas T60 F1 under foliar application of melatonin showed 6.02 % increase in number of viable seeds per fruit (93.90) as compared to control (88.28).
Similarly in the case of heat-sensitive genotypes of tomato a significant increase (3.72 %) in number of viable seeds per fruit was observed in Naqeeb (83.78) as compared to the control (80.77). While Nagina under foliar application of melatonin presented the 4.49 % more number of viable seeds per fruit (81.45) as compared to the control (77.95).
3.3 Impact of melatonin foliar treatment on physiological traits of tomato genotypes
Data regarding photosynthetic rate (Fig. 1) reflected that 42.04 % increase in photosynthesis rate from heat tolerant genotypes of tomato was observed in Super cash F1 (28.14 μmol CO2 m−2 S-1) as compared to without application of melatonin i.e. control (19.81 μmol CO2 m−2 S-1). However, T60 F1 under foliar application of melatonin showed 39.95 % increase in photosynthesis rate (26.38 μmol CO2 m−2 S-1) as compared to control (18.85 μmol CO2 m−2 S-1).![Foliar application of melatonin improved Physiological [Photosynthetic rate (a), Transpiration rate (b), Stomatal conductance (c), SPAD Chlorophyll contents (d)] in heat tolerant and heat sensitive genotypes of tomato under heat stress. Mean value ± standard error, significant difference is exhibited by lower case letters (p ≤ 0.05) according to LSD.](/content/185/2024/36/10/img/10.1016_j.jksus.2024.103420-fig1.png)
Foliar application of melatonin improved Physiological [Photosynthetic rate (a), Transpiration rate (b), Stomatal conductance (c), SPAD Chlorophyll contents (d)] in heat tolerant and heat sensitive genotypes of tomato under heat stress. Mean value ± standard error, significant difference is exhibited by lower case letters (p ≤ 0.05) according to LSD.
In case of heat-sensitive genotypes of tomatoes, the greater photosynthesis rate was observed in Naqeeb (21.64 μmol CO2 m−2 S-1) as compared to control (16.26 μmol CO2 m−2 S-1) indicating 33.08 % improvement over control. Whereas, Nagina under foliar application of melatonin presented 31.73 % improvement in photosynthesis rate (18.39 μmol CO2 m−2 S-1) as compared to its respective control (13.96 μmol CO2 m−2 S-1). The foliar treatment of melatonin, statistically (p ≤ 0.05) increased the photosynthesis rate in heat tolerant as well as in heat sensitive cultiars of tomato.
In the same way, significant (p ≤ 0.05) results of melatonin foliar spray swere obtained in case of transpiration rate (Fig. 1). According to the results, maximum transpiration rate was noted in Super cash F1 (2.31 mmol H2O m−2 S-1) as compared to control (1.78 mmol H2O m−2 S-1) reflecting 29.77 % increase over control. Similarly, T60 presented 27.93 % more transpiration rate (2.29 mmol H2O m−2 S-1) as compared to its respective control (1.79 mmol H2O m−2 S-1). Similarly, in heat-sensitive genotypes of tomato, results expressed that the 23.27 % increase in transpiration rate was observed in Naqeeb (1.96 mmol H2O m−2 S-1) as compared to control (1.59 mmol H2O m−2 S-1). While Nagina reflected minimum increase (23.53 %) in transpiration rate (1.89 mmol H2O m−2 S-1) than its control (1.53 mmol H2O m−2 S-1).
The results (Fig. 1) indicated that under the foliar application of melatonin improved 26.63 % stomatal conductance significantly (p ≤ 0.05) in T60 F1 (2.33 mmol m−2 S-1) than its control (1.84 mmol m−2 S-1). While Super cash F1 genotype showed 27.93 % improvement in stomatal conductance (2.29 mmol m−2 S-1) as compared to its respective control (1.79 mmol m−2 S-1). Similarly, heat-sensitive genotypes of tomatoes also revealed the same trend as heat-tolerant genotypes.
It is clear from the data (Fig. 1) regarding chlorophyll contents that foliar application of melatonin increased chlorophyll contents significantly (p ≤ 0.05) in all tomato cultivars. The results showed that under the foliar application of melatonin greater chlorophyll contents from heat-tolerant genotypes of tomato were observed in T60 F1 (29.04 mg g−1) as compared to control (21.23 mg g−1) untreated showing 36.78 % increase in chlorophyll contents. Super cash F1 under foliar application of melatonin indicated 39 % improvement in chlorophyll contents (28.12 mg g−1) as compared to its respective control (20.23 mg g−1).
In the case of heat-sensitive genotypes of tomato, the results revealed that foliar application of melatonin increase 26.95 % chlorophyll contents in Naqeeb (21.86 mg g−1) as compared to control (17.22 mg g−1). Whereas, Nagina presented 25.16 % more chlorophyll contents (21.19 mg g−1) as compared to control (16.93 mg g−1).
The results (Fig. 2) regarding WUE had revealed that the highest increase in WUE (9.43 %) was noted in the Super cash F1 (12.18 μmol CO2/ mmol H2O) genotype as compared to the control (11.13 μmol CO2/ mmol H2O). In the same way, T60 F1 under foliar application of melatonin presented 9.41 % increase in WUE (11.51 μmol CO2/ mmol H2O) as compared to its control (10.52 μmol CO2/ mmol H2O). Similar to heat-tolerant genotypes, 8.03 % improvement in WUE in heat-sensitive genotypes of tomato was observed in Naqeeb (11.03 μmol CO2/ mmol H2O) as compared to control (10.21 μmol CO2/ mmol H2O). Subsequently, Nagina under foliar application of melatonin showed greater WUE (9.71 μmol CO2/ mmol H2O) than its respective control (9.10 μmol CO2/ mmol H2O) indicating 6.70 % increase over control.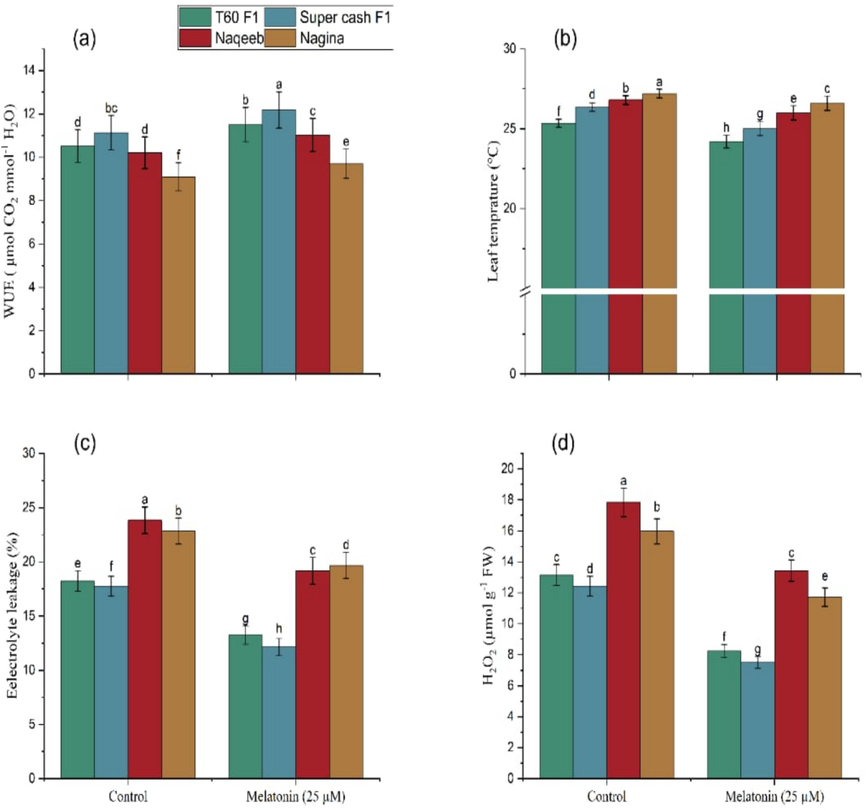
Foliar application of melatonin improved Water use efficiency (WUE) (a), Leaf temperature (b), and reduced Electrolyte leakage (%) (c), Hydrogen per oxide (H2O2) (d) in heat tolerant and heat sensitive genotypes of tomato under heat stress. Mean value ± standard error, significant difference is exhibited by lower case letters (p ≤ 0.05) according to LSD.
The leaf temperature (Fig. 2) of heat-tolerant genotype T60 F1 (24.20 °C) showed 4.53 % low leaf temperature as compared to control (25.35 °C). Leaf temperature of other heat tolerant genotypes Super cash F1 was also reduced (25 °C) as 5.12 % over its control (26.35 °C). On the other hand, 2.20 % lower leaf temperature in heat-sensitive genotypes of tomato in Nagina (26.60 °C) was observed as compared to control (27.20 °C). Whereas, another heat-sensitive cultivar Naqeeb also showed 2.98 % lower leaf temperature (26 °C) than its respective control (26.80 °C).
The results regarding electrolyte leakage (EL) indicated that the minimum EL was recorded in Super cash F1 (12.15 %) as compared to control (17.76 %). Similarly, T60 F1 under foliar application of melatonin presented the lowest electrolyte leakage (13.24 %) as compared to control (18.23 %). Similar to heat-tolerant genotypes of tomato foliar application of melatonin also decreased the electrolyte leakage in heat-sensitive genotypes of tomato and results expressed that the lowest electrolyte leakage was observed in Nagina (18.67 %) than its control (22.83 %). Whereas, Naqeeb also showed minimum electrolyte leakage (19.15 %) than its control (23.83 %).
3.4 Impact of melatonin foliar treatment on ROS of tomato genotypes
The maximum reduction (13.15 %) in H2O2 contents (Fig. 2) under the foliar application of melatonin from heat tolerant genotypes of tomato was noted in T60 F1 (8.24 µmol g−1 FW) as compared to without application of melatonin (13.15 µmol g−1 FW). Similarly, in another heat tolerant cultivar Super cash F1 also presented 39.50 % reduction in H2O2 contents (7.52 µmol g−1 FW) than its respective control (12.43 µmol g−1 FW). In case of heat sensitive genotypes of tomato,24.72 % reduction in H2O2 contents were observed in Naqeeb (13.43 µmol g−1 FW) when compared with control (17.84 µmol g−1 FW) and followed by Nagina showing the 26.61 % less reduction in H2O2 (11.72 μmol g−1 FW) as compared to its control (15.97 µmol g−1 FW).
3.5 Impact of foliar melatonin treatment on leaf and root osmoprotectants (GB and proline) of tomato genotypes
The maximum increase in GB contents (Fig. 3) in leaf and roots of heat-tolerant genotypes of tomato was noted in T60 F1 (22.76 % and 17.14 %) in leaf and roots respectively as compared to without application of melatonin. Similarly, in another heat-tolerant cultivar Super cash F1 also presented 20 % and 15.53 % higher GB contents in leaf and roots than its respective control. In the case of heat-sensitive genotypes of tomato, 13.33 % and 9.52 % more GB contents in leaf and roots were observed in Naqeeb as compared to the control followed by Nagina (12.78 % and 9.30 %) showed high GB contents in leaf and roots respectively as compared to their respective control.![Foliar application of melatonin improved osmoprotectants [Leaf GB (a), Root GB (b), Leaf proline (c), Root proline (d)] in heat tolerant and heat sensitive genotypes of tomato under heat stress. Mean value ± standard error, significant difference is exhibited by lower case letters (p ≤ 0.05) according to LSD.](/content/185/2024/36/10/img/10.1016_j.jksus.2024.103420-fig3.png)
Foliar application of melatonin improved osmoprotectants [Leaf GB (a), Root GB (b), Leaf proline (c), Root proline (d)] in heat tolerant and heat sensitive genotypes of tomato under heat stress. Mean value ± standard error, significant difference is exhibited by lower case letters (p ≤ 0.05) according to LSD.
The maximum increase in proline contents (Fig. 3) in leaf and roots of heat tolerant genotypes of tomato was noted in T60 F1 (20.95 % and 15.05 %) respectively as compared to without application of melatonin. Similarly, in other heat-tolerant cultivar Super cash F1 also presented 20.88 % and 14.28 % higher proline contents in leaf and roots respectively than their respective control. In case of heat-sensitive genotypes of tomato, maximum increase in proline contents of leaf (13.28 %) and roots (10.81 %) were observed in Naqeeb as compared to the control followed by Nagina showed 13.70 % and 11.26 % increase in proline contents in leaf and roots respectively as compared to their respective control.
3.6 Impact of melatonin foliar treatment on enzymatic antioxidant activities of tomato genotypes
The enzymatic antioxidants activities (NOS, NR, GR, and GST) in all genotypes (heat tolerant and heat sensitive) of tomato were significantly (p ≤ 0.05) improved (Fig. 4) when melatonin was applied as compared to control which remained under heat stress without melatonin treatment. Maximum increase in activity of NOS (133.91 %), NR (48.38 %), GR (59.76 %) and GST (55.92 %) was found in heat tolerant genotype (Super cash F1) under heat stress along with melatonin foliar spray. However, both heat tolerant cultivars of tomato showed a non-significant difference in increasing activity of enzymes. while minimum activity of enzymes were indicated by heat sensitive genotypes (Naqeeb and Nagina) as compared to control treatment which remained untreated bearing heat stress.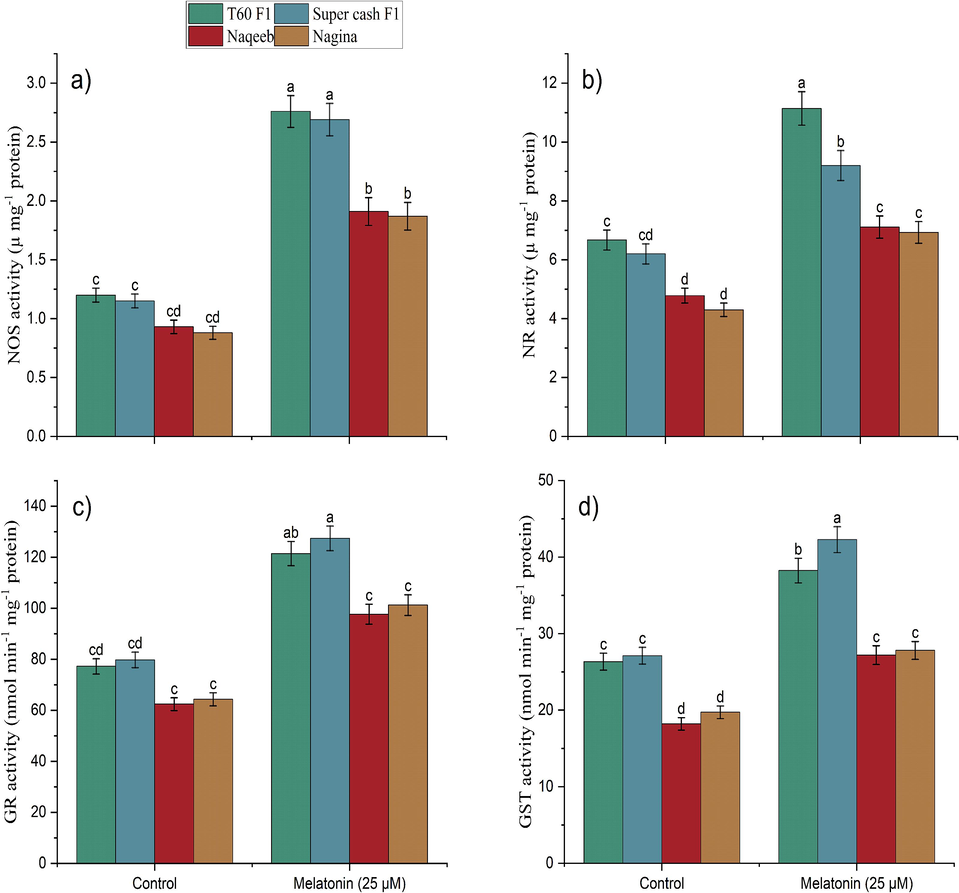
Foliar application of melatonin improved Nitric Oxide Synthase (NOS) activity (a), Nitrate reductase (NR) activity(b), glutathione reductase (GR) activity(c), glutathione S-transferase (GST) activity (d) in heat tolerant and heat sensitive genotypes of tomato under heat stress. Mean value ± standard error, significant difference is exhibited by lower case letters (p ≤ 0.05) according to LSD.
3.7 Pearson association between morphological, physiological, quality and enzymatic traits of tomato genotypes
The Pearson correlation analysis revealed that there was a significant relationship between morphological, physiological, quality and enzymatic traits of tomato genotypes (Fig. 5). Moreover, the photosynthetic rate, transpiration rate, stomatal contents, chlorophyll contents, WUE, GB, proline, NOS, NR, GR, GST, plant fresh biomass, plant dry biomass, leaf area, stem girth, protein contents, and number of viable seeds were positively associated with each other (p ≤ 0.05) and negatively associated with the EL, leaf temperature, and H2O2.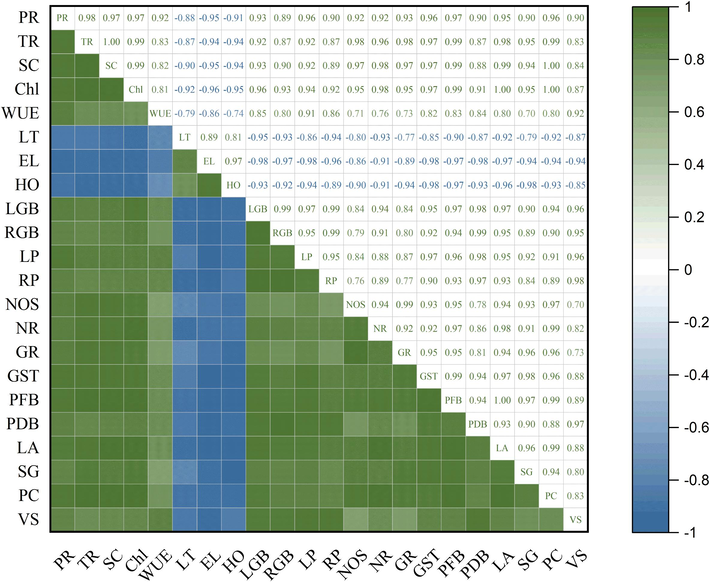
Correlation analysis of Photosynthetic rate (PR), Transpiration rate (TR), Stomatal conductance (SC), SPAD chlorophyll contents (Chl), Water use efficiency (WUE), Leaf temperature (LT), reduced Electrolyte leakage (EL), Hydrogen per oxide (HO), Leaf GB, Root GB, Leaf proline (LP), Root proline (RP), Nitric oxide synthase (NOS) activity, Nitrate reductase (NR) activity, Glutathione reductase (GR) activity, Glutathione S-transferase (GST) activity, Plant fresh biomass (PFB), Plant dry biomass(PDM), Leaf area (LA), Stem grith (SG), Protein contents (PC), Viable seeds (VS) in heat tolerant and heat sensitive genotypes of tomato under heat stress.
4 Discussion
Melatonin is a signaling molecule known as a pleiotropic molecule which is capable of improving heat tolerance in plants by alleviating its adverse effects. Numerous researches have explored its determined role in improving plant physiology by regulating growth mechanisms. However, the comprehensive role of melatonin in improving plant growth and yield attributes under abiotic stress conditions is not yet determined. Therefore to understand its role in the mitigation of heat stress foliar spray was applied to four genotypes of tomatoes including heat-tolerant and heat-sensitive cultivars.
A foliar spray of melatonin increased fresh and dry biomass in both heat-tolerant and heat-sensitive genotypes of tomatoes under heat-stress conditions. The reduction in fresh biomass and dry biomass of plants is directly related to heat stress. High temperature disrupted the water potential owing to increased ROS species leading to reduced photosynthetic activity, chlorophyll contents, and water use efficiency (Wang et al., 2022). All these processes are responsible for lowering of fresh and dry biomass of plants. However foliar application of melatonin influenced all these growth attributes positively and increased plant fresh and dry biomass by stabilizing water potential and enzymatic activities to reduce ROS species that are responsible for membrane stability (Ahmad et al., 2023). Melatonin also acts as a growth promoter and induces the activity of auxins and indole acetic acid (IAA) that contribute to improved plant vegetative growth and cell expansion in plants which subsequently increases plant biomass (Arnao and Hernández‐Ruiz, 2021).
Stem grith of both heat tolerant and heat sensitive genotypes of tomato reduced under heat stress. It is also directly related to water loss in high amounts from plants. High water loss reduced water use efficiency in plant metabolism by inducing oxidative damage which subsequently reduced plant growth in terms of plant growth (Kapoor et al., 2020). Melatonin spray increases plant growth by increasing water uptake due to low oxidative damage of the membrane (Nawaz et al., 2020). The leaf area is also reduced under heat shock. Under high-temperature shock, plants experience low water potential due to which uptake of water and nutrients become restricted. Thus limited water retention in plant metabolism caused stunted growth of plants and yellowing of leaves due to leaf senescence. Thus, high water loss under heat stress also contributed to reducing leaf area by Leaf senescence. Melatonin spray influences leaf area in a positive manner by mitigating this adverse effect of heat stress and increasing heat stress tolerance by stabilizing reactive oxygen species (ROS) and photosynthetic electron flux which is responsible for increased Fv/Fm ratio in plants (Altaf et al., 2022).
Heat stress also contributed to depressing quality of tomatoes in terms of low protein contents and poor viability of seeds in both heat tolerant and heat sensitive cultivars of tomato. Melatonin foliar spray improved protein contents in tomatoes owing to its contribution in increasing the activity of antioxidant enzymes which acted as scavengers for reactive oxygen species (ROS) leading to reduced misfolding and denaturing of protein as well as started to refold denatured protein (Hassan et al., 2022). Similarly, Poor viability in seeds was induced due to pollen abortion is caused by heat stress (Lohani et al., 2022). melatonin foliar spray increased the viability and number of seeds in all tomato cultivars. This might be due to the pleiotropic nature of melatonin which acts as a phyto-regulator to promote the development and growth of plants during the reproductive stage and plays its regulatory role in pollen thermotolerance in all genotypes of tomato (Colombage et al., 2023).
Photosynthetic rate, chlorophyll contents, stomatal conductance, and transpiration rate are the physiological processes that directly depend upon each other (Jaffar et al., 2023; Sadaf et al., 2023). Improvement of one attribute will improve another. Regulation of all these physiological attributes declined under heat shock due to restricted water and nutrient availability. Under low water uptake and high temperature, these physiological processes deteriorated due to the accumulation of oxidative stress and membrane damage which subsequently caused disturbance in balanced homeostasis (Tiwari et al., 2020). Melatonin acts as a growth regulator and improves photosynthetic rate by improving photosynthetic efficiency, chlorophyll contents, stomatal conductance, and transpiration rate by improving the activity of antioxidant enzymes to minimize oxidative stress (Hassan et al., 2022; Hasan et al., 2023). Meanwhile, heat stress also caused the induction of chlorophyllase enzymes which reduced chlorophyll contents. Melatonin has the capability to reduce the activity of this enzyme and to promote the synthesis of chlorophyll pigment (Javed et al., 2022). However, melatonin spray regulates the opening of stomata for regular exchange of gases by improving membrane turgidity. Last but not least melatonin foliar spray increased all physiological parameters of both heat-tolerant and heat-sensitive genotypes of tomato (Annadurai et al., 2023).
Heat stress also influenced water use efficiency (WUE) negatively. High temperature restricted water uptake due to which regulation of opening and closing of stomata distorted lead to reduced WUE by losing turgidity of the membrane. Melatonin improves water use efficiency by maintaining water potential within plants to regulate membrane turgidity by the accumulation of soluble compounds and by reducing membrane damage owing to ROS oxygen species (Jahan et al., 2021). High temperature induced electrolyte leakage (EL), leaf temperature, and hydrogen peroxide (H2O2) due to oxidative stress. To make a conducive environment for plant growth under heat stress (Barman et al., 2019; Annadurai et al., 2023), soluble solutes or osmoprotectants such as proline and glycine betaine (GB) were increased in both leaf and roots under heat stress. A foliar spray of melatonin further increased these osmoprotectants to protect plants from the negative effects of heat stress. Melatonin reduced ROS species to induce a strong defense system and increased regulation of proline and GB within the plant by maintaining water requirements leading to reduced osmotic stress (Mushtaq et al., 2022). These compatible solutes also improve nitrogenous compounds within plants' regulated nutrient supply (Alharbi et al., 2021).
Melatonin foliar spray augmented the regulation of enzymatic antioxidant like Nitric oxide synthase (NOS), Glutathione reductase (GR), nitrate reductase (NR) and glutathione S-transferase (GST)) to make a strong defense system of plants against oxidative stress (Awan et al., 2023). Under heat stress, melatonin provide a condusive environment to enzymes activity. This increase in activity might be attributed to hormonal regulation caused by melatonin which influenced hormonal balance and signal transduction which activated signal pathways to promote antioxidant activities leading to protect plant from heat stress. Moreover, melatonin also directly acts as antioxidant and protects the plants from oxidative damage (Hassan et al., 2022). The antioxidants act as scavenger of ROS thus improving the physiological and morphological traits of tomato plants (Javed et al., 2022; Khan et al., 2024). Infact under abiotic stress (control) activity of GR, NR, NOS and GST were downregulated owing to declined expression of genes related to each enzyme that were improved in both genotypes of tomato when melatonin was applied in the form of foliar spray (Jahan et al., 2019).
5 Conclusion
The present study explored the adverse effect of heat stress due to low water uptake and high temperature. Heat shock causes stunted growth of plants owing to oxidative damage, impaired photosynthetic efficiency, and high ROS species. To improve heat tolerance melatonin spray was applied to the tomato cultivars. Melatonin acts as a growth regulator and provides a conducive environment for plant growth under heat stress by improving the growth and physiological attributes of tomato plants in both heat-tolerant and heat-sensitive genotypes. Melatonin also improves antioxidant enzymetic activity (NOS, NR, GR, GST) to minimize oxidative stress and restore membrane turgidity under heat-stress environment. Moreover, protein contents and viability of tomato seeds were also recovered by melatonin foliar spray due to ROS-mediated damage. This study was confined to pot experiments and in the climatic region of Sargodha. It is required to perform this experiment in future in all ecological zones under field conditions to understand the interactive effect of heat stress and melatonin foliar spray in depth.
Short title: Melatonin Foliar Spray Mitigates Heat Stress in Tomato Cultivars.
Ethics approval.
Not applicable.
Consent to participate.
All authors consent to participate in the manuscript publication
Consent for publication
All authors approved the manuscript to be published.
CRediT authorship contribution statement
Hafiz Muhammad Tayyab Khan: Visualization, Methodology, Investigation, Conceptualization. Syed Ayyaz Javed: Methodology, Conceptualization. Muhammad Tauseef Jaffar: Methodology, Conceptualization. Rashad Mukhtar Balal: Methodology, Conceptualization. Qurat ul Ain: Writing – original draft, Data curation, Conceptualization. Alaiha Asif: Writing – original draft, Data curation, Conceptualization. Muhammad Adnan Shahid: Writing – original draft, Data curation, Conceptualization. Mohamed A. El-Sheikh: Writing – review & editing, Conceptualization. Parvaiz Ahmad: Writing – review & editing, Conceptualization.
Acknowledgements
The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2024R182) King Saud University, Riyadh, Saudi Arabia.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Mechanisms of elevated CO2-induced thermotolerance in plants: the role of phytohormones. Plant Cell Reports. 2021;40(12):2273-2286.
- [Google Scholar]
- Melatonin-induced detoxification of organic pollutants and alleviation of phytotoxicity in selected horticultural crops. Horticulturae. 2022;8(12):1142.
- [Google Scholar]
- The role of melatonin in plant growth and metabolism, and its interplay with nitric oxide and auxin in plants under different types of abiotic stress. Front. Plant Sci. 2023;14:1108507.
- [Google Scholar]
- Exogenous application of melatonin alleviates salt stress-induced decline in growth and photosynthesis in Glycine max (L.) seedlings by improving mineral uptake, antioxidant and glyoxalase system. Plant Soil Environ. 2021;67:208-220.
- [Google Scholar]
- Melatonin improves drought stress tolerance of tomato by modulating plant growth, root architecture, photosynthesis, and antioxidant defense system. Antioxid. 2022;11:309.
- [Google Scholar]
- Melatonin Decreases Negative Effects of Combined Drought and High Temperature Stresses through Enhanced Antioxidant Defense System in Tomato Leaves. J. Hortic. 2023;9:673.
- [Google Scholar]
- A review of heat stress signaling in plants. IOP Conference Series: Earth and Environ. Sci. 2020;484:012041
- [Google Scholar]
- Melatonin as a regulatory hub of plant hormone levels and action in stress situations. Plant Biol. 2021;23:7-19.
- [Google Scholar]
- Exogenous Melatonin Regulates the Expression Pattern of Antioxidant-Responsive Genes, Antioxidant Enzyme Activities, and Physio-chemical Traits in Pearl Millet Under Drought Stress. Journal of Plant Growth Regulation 2023:1-15.
- [Google Scholar]
- Amelioration of heat stress during reproductive stage in rice by melatonin. Indian J. Agric. Sci. 2019;89:1151-1156.
- [Google Scholar]
- Microtubule Organization at the Cell Cortex is a Determinant of Cell Shape via Division Plane Maintenance and Directional Cell Expansion in Plants. Riverside: University of California; 2020.
- A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 1976;72:248-254.
- [Google Scholar]
- Involvement of hydrogen peroxide in the regulation of senescence in pear. Plant Physiology. 1977;59:411-416.
- [Google Scholar]
- Melatonin and abiotic stress tolerance in crop plants. Intern. J. Mole. Sci. 2023;24:7447.
- [Google Scholar]
- Fahad, S., Adnan, M., Hassan, S., Saud, S., Hussain, S., Wu, C.H., J, 2019. Rice responses and tolerance to high temperature. In: Advances in rice research for abiotic stress tolerance, Woodhead Publishing, pp: 201–224.
- The potential of resilient carbon dynamics for stabilizing crop reproductive development and productivity during heat stress. Plant Cell & Environ. 2021;44:2066-2089.
- [Google Scholar]
- Statistical Procedures for Agricultural Research (2nd ed.John 430). New York: Wiley and Sons; 1984.
- Melatonin mediates elevated carbon dioxide-induced photosynthesis and thermotolerance in tomato. Journal of Pineal Research. 2023;74(3):e12858.
- [Google Scholar]
- Melatonin: a vital pro-tectant for crops against heat stress: mechanisms and prospects. J. Agron. 2022;12
- [Google Scholar]
- Physiology of heat stress and tolerance mechanism—an overview. J. Plant Sci Res. 2018;33:55-68.
- [Google Scholar]
- Exogenous melatonin induces drought stress tolerance by promoting plant growth and antioxidant defence system of soybean plants. AoB Plants. 2021;13:026.
- [Google Scholar]
- Beneficial interaction of allelopathic bacteria with chemical herbicides for sustainable wheat (Triticum aestivum L.) production under wild oat (Avena fatua L.) infestation. Italian. J. Agron. 2023;18
- [Google Scholar]
- Melatonin alleviates heat-induced damage of tomato seedlings by balancing redox homeostasis and modulating polyamine and nitric oxide biosynthesis. BMC Plant Biol. 2019;19:1-16.
- [Google Scholar]
- Melatonin-mediated photosynthetic performance of tomato seedlings under high-temperature stress. Plant Physiol. Biochem. 2021;167:309-320.
- [Google Scholar]
- Interactive effect of different salinity sources and their formulations on plant growth, ionic homeostasis and seed quality of maize. Chemo. 2022;291:132678
- [Google Scholar]
- Protective stabilization of mitochondrial permeability transition and mitochondrial oxidation during mitochondrial Ca2+ stress by melatonin's cascade metabolites C3-OHM and AFMK in RBA1 astrocytes. J. Pineal Res. 2019;66:12538.
- [Google Scholar]
- The impact of drought in plant metabolism: How to exploit tolerance mechanisms to increase crop production. Applied Sci. 2020;10:5692.
- [Google Scholar]
- Exogenous application of melatonin mitigate the heat stress in different tomato (Solanum lycopersicum L.) cultivars. J. King Saud University-Sci. 2024;36:103086
- [Google Scholar]
- Short-term heat stress during flowering results in a decline in Canola seed productivity. J. Agron. Crop Sci. 2022;208:486-496.
- [Google Scholar]
- Separation of hydrogen peroxide from organic hydroperoxides. Analytical Chemistry. 1953;25:1760-1761.
- [Google Scholar]
- Melatonin in micro-tom tomato: Improved drought tolerance via the regulation of the photosynthetic apparatus, membrane stability, osmoprotectants, and root system. Life. 2022;12
- [Google Scholar]
- Melatonin as master regulator in plant growth, development and stress alleviator for sustainable agricultural production: current status and future perspectives. Sustain. 2020;13:294.
- [Google Scholar]
- Impact of drought and heat stress individually and in combination on physio-biochemical parameters, antioxidant responses, and gene expression in Solanum lycopersicum. 3 Biotech. 2020;10:1-18.
- [Google Scholar]
- Rivero, R.M., M, K., A, G., H, S., R, M., S, G., E, B., 2007. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. pp: 19631–19636.
- Influence of brassinosteroid and silicon on growth, antioxidant enzymes, and metal uptake of leafy vegetables under wastewater irrigation. Int. J 2023:1-11.
- [Google Scholar]
- Shafi, A., Singh, A.K., Zahoor, I., 2021. Melatonin: Role in abiotic stress resistance and tolerance. In: Plant growth regulators: signalling under stress conditions, pp: 239–273.
- Effects of different treatments of salicylic acid on heat tolerance, chlorophyll fluorescence, and antioxidant enzyme activity in seedlings of Cucumis sativa L. Plant Growth Regul. 2006;48:127-135.
- [Google Scholar]
- Melatonin as a potent and inducible endogenous antioxidant: Synthesis and metabolism. Mol. 2015;20:18886-18906.
- [Google Scholar]
- Tiwari, S., Patel, A., Singh, M., Prasad, S.M., 2020. Regulation of temperature stress in plants. In: Plant life under changing environment, Academic Press, pp: 25–45.
- Heat shock proteins: dynamic biomolecules to counter plant biotic and abiotic stresses. Int. J. Mol. Sci. 2019;20:5321.
- [Google Scholar]
- Linking plants functioning to adaptive responses under heat stress conditions: a mechanistic review. J. Plant Growth Regul 2021:1-18.
- [Google Scholar]
- Effects of high-temperature stress on photosynthetic characteristics and antioxidant enzyme system of Paeonia ostii. Phyton. 2022;91:599.
- [Google Scholar]
Appendix A
Supplementary data
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jksus.2024.103420.
Appendix A
Supplementary data
The following are the Supplementary data to this article:




