

Translate this page into:
Using two-sex life table tools to compare the population parameters of Oxycarenus hyalinipennis costa (Lygaeidae: Hemiptera) when fed on Bt and non-Bt cotton seeds
⁎Corresponding author. waqar4me@yahoo.com (Waqar Jaleel)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Cotton crop (Gossypium hirsutum L.) is considered economy crop worldwide especially in Pakistan. Cotton plays strong role in proportion of GDP and foreign exchange earnings. In Pakistan, around 55% cotton is exporting fiber commodity and 15 % area of cultivation is devoted to cotton crop especially Bt cotton. However, Bt cotton suppress the chewing pests but give a favorable niche to sucking pests. Among sucking pests, the Oxycarenus hyalinipennis Costa is emerging sucking pest in Bt cotton, and damaging the more than 40 different plants. Study regarding preference of seed and two-sex life table traits of O. hyalinipennis is necessary for its eco-friendly management. Therefore, this explain the preference of cotton seeds e.g., CIM-620 (non-Bt variety), CIM-717 (non-Bt variety), Bt-CIM-632 and Bt-Cyto-179 by O. hyalinipennis, as well as its fitness on these seeds under laboratory condition. Results indicated that both males and females preferred Bt-Cyto-179 for ovipositing as compared to other three varieties. The developmental time from eggs to males (176.85 ± 7.43) and females (184.94 ± 7.85) were significantly longer on other verities as compared to Bt-Cyto-179. The O. hyalinipennis females produce significantly more eggs on Bt-Cyto-179 (215 ± 2.34 eggs female-1) than those on other three seeds e.g., CIM-620, CIM 717, and Bt-CIM-632. In conclusion, Bt-Cyto-179 was the more suitable varieties to attract and rear O. hyalinipennis than CIM 632, and CIM 717 seeds. And our study suggests that Bt-Cyto-179 can be used as a bait crop to control O. hyalinipennis and rearing purpose for molecular studies.
Keywords
Oxycarenus hyalinipennis
Cotton Seeds
Seed Preference
Bt and non Bt varieties

1 Introduction
Cotton, Gossypium hirsutum L., especially Bt-cotton is considered economical crop and provide major income source to people all over the world especially in Pakistan (Ahmad et al., 2020; Arshad et al., 2021). Last few years, the area of Bt-cotton cultivation decreased in Pakistan because of various factors that affecting the cotton productivity in Pakistan (Arshad et al., 2021; Khan et al., 2014a), as one of insect pest because cotton crop is more susceptible nature. However, previous literatures proved that Bt cotton suppressed bollworms but found less effective against sucking pests except a few thrips species (Jaleel et al., 2014; Razaq et al., 2019). Another reason is unselective and blind application of insecticides affecting the environmentally safety (Poudel et al., 2020); moreover, it also caused resistance in insects that reduced their vulnerability to the insecticides (Jaleel et al., 2020; Lei et al., 2021; Saeed et al., 2016). However, nowadays in Pakistan, among sucking pest of cotton, the Oxycarenus hyalinipennis Kirby (Lygaeidae: Hemiptera) is considered damaging bug in cotton crop.
Now, the O. hyalinipennis has gained the status of major pests of cotton crop in Pakistan, as well as in other countries e.g., China, Turks, Islands of U.S. Bahamas, Caicos, and Cayman (Colmenárez et al., 2020; Stocks and Peña, 2013). Although it is a well-known pest of crops of family Malvaceae, its infestation has been observed on other crops like guava, moringa, mango, okra, chilies and lemon (Naeem-Ullah et al., 2020). It’s adults as well as nymphs are responsible for affecting the quality and quantity of yield and oil extraction of cotton seeds. Its adults and nymphs sociably feeds on squares of cotton plant, resulting in pale yellow color of squares and then shedding of infested squares (Rajendran et al., 2018). Lot of factors may affect the population parameters of dusky cotton bug e.g. adaptation of novel techniques in cotton crop like early sowing of Bt-cotton and extreme use of insecticides (Khan and Ghramh, 2021a; Rocha-Munive et al., 2018). Such factors are creating niche for a survival of dusky cotton bug and this insect pest is gaining the status of economically important pests in Pakistan (Khan and Ghramh, 2021b; Naeem-Ullah et al., 2020).
Use of insecticides is considered a reliable method against this insect pest in Pakistan and other developing countries (Ghramh et al., 2020; Hashmi and Khan, 2011). But excessive use of insecticides causes serious environmental and health problems for human being and non-target organisms. Frequent use of insecticides also causes the problems of resistance development in insects (Ghramh et al., 2019; Khan et al., 2014b). Dusky cotton bug has also developed resistance against different group of insecticides (Banazeer et al., 2021). However, the alternative control strategy is needed for the management of dusky cotton bug.
Basic information of insect pests especially O. hyalinipennis are important in the integrated management program (Ijaz and Shad, 2021a; Jaleel et al., 2018; Sarmad et al., 2020). However, the fitness of bugs can be better explained by using two-sex life table tools to find the susceptible point (Huang and Chi, 2012). The study regarding life table parameters of O. hyalinipennis have been explained on different hosts including cotton (Ananthakrishnan et al., 1982; Banazeer et al., 2021; Ijaz and Shad, 2021b; Wazir and Shad, 2021); however, few studies are available on cotton seeds preference by O. hyalinipennis and its fitness. Considering the importance of cotton crop this research explains the behaviour of O. hyalinipennis on two non-Bt (CIM 620 and CIM 717) and Bt (Bt-CIM-632 and Bt-Cyto-179) varieties in the laboratory.
2 Materials and methods
2.1 Insect collection, rearing, and seeds
The O. hyalinipennis was collected from different cotton fields and maintained in laboratory of Entomology, Central Cotton Research Institute (CCRI), Multan, Punjab, Pakistan. Dusky cotton bugs were shifted into jugs having dimension 14 × 24 cm. Temperature (26 ± 2 °C) and humidity (60 ± 5%) were maintained in laboratory using air conditioner (Haier Pear 18HFAA, Ind. Ltd Pk) and humidifier (Life Care 600 Ind. Ltd Pk) with photoperiod 15: 9 h L: D. The O. hyalinipennis were fed on soaked seed of each variety. Population of dusky cotton bugs were reared on each seed e.g., two non-Bt (CIM 620 and CIM 717) and Bt (Bt-CIM-632 and Bt-Cyto-179) for two generation to maintain homogenous population and make them suitable for laboratory study.
2.2 Data recording
After populations of O. hyalinipennis acclimatized, the two sex life table parameters were studied on two non-Bt (CIM 620 and CIM 717) and Bt (Bt-CIM-632 and Bt-Cyto-179) seeds. Hundred eggs of dusky cotton bugs on each seed were collected and shifted on respective seeds in 14 × 24 cm plastic jars to record the two-sex life table parameters. After adult formation, A pairs (male: female) of adults shifted into a Petri dish (9 × 1.5 cm) for fecundity. Total 10 replication was done on each cotton seeds.
2.3 Protein analysis
Buffers were used to maintain enzyme nature during crushing of tested seeds. The ingredients for extracting protein were used as distilled water, Na2HPO412H2O, NaH2PO42H2O, blue dye, phosphoric acid, and ethanol. As first 8.10 g of Na2HPO42H2O crystals was poured into volumetric flask having water 250 ml and we designated as “A”. And 0.35 g of Na2HPO42H2O was dissolved in 200 ml of water and we designated as “B”. Then, 20 ml solution of “A” and 80 ml solution of “B” were mixed for maintained pH 7. This solution was used to find out the protein values in seed contents.
2.4 Statistical analysis
The statistics software was used to calculate the mean and the LSD test variation among seeds treatments. The two-sex life table traits (Chi, 1988; Chi and Liu, 1985) were calculated using TWO-SEX LIFE TABLE program (Chi, 1988; Liu et al., 2017). The values of population and age stage parameters R0, and sxj, fxj, lx, mx, exj, vxj respectively were calculated as described in the methodology of (Chi, 1988; Jaleel et al., 2018). The following equations explained the two-sex life table calculations.
3 Results
3.1 Life history parameters
Longevity of different nymphal instars on different seeds was significantly different. Larval instars’ duration increases when reared on Bt-Cyto-179 (Table 1). First larval instar developed longer on Bt-Cyto-179 than other tested seeds (P < 0.0001). Similarly, same pattern was observed in other instars. Adults of O. hyalinipennis lived longer on Bt-Cyto-179 (P < 0.0001). The APOP period was significantly longest on CIM 632 (P < 0.001). While, maximum TPOP duration was recorded on Bt-Cyto-179 seeds as compared to other three seeds (P < 0.0001) (Table 1). Number of eggs increased dynamically as to optimum condition/maximum numbers achieved on Bt-Cyto-179 (Table 1). Fecundity was significantly highest when dusky cotton bugs fed on Bt-Cyto-179 seeds as compared to three other seeds (P < 0.0001) (Table 1). n: represent the no. of individual, APOP: Adult pre-oviposition period of a female adult, APRP: Adult pre-reproduction period of a female adult, The standard errors of the mean values of pre-adult development period and total longevity of male and females were estimated by using 200,000 bootstrap replicates. Means marked with different letters are significantly different between three treatments using the paired bootstrap test at the 5% significant level.
Cotton Seeds
Non-Bt Bt
Parameters
CIM 620
CIM 717
Bt-Cyto-179
CIM 632
Egg (d)
5.06 ± 0.19b (n: 100)
4.62 ± 0.23c (n:100)
6.37 ± 0.18a (n:100)
5.09 ± 0.23b (n:100)
1st instar (d)
4.16 ± 0.18c (n: 74)
5.39 ± 0.29b (n:66)
5.58 ± 0.30a (n:93)
4.05 ± 0.23d (n:66)
2nd instar (d)
4.12 ± 0.29b (n: 50)
4.38 ± 0.29a (n:47)
3.61 ± 0.29c (n:72)
4.17 ± 0.25b (n:47)
3rd instar (d)
3.21 ± 0.09d (n: 47)
3.94 ± 0.15b (n:40)
3.58 ± 0.11c (n:60)
4.08 ± 0.11a (n:40)
4th instar (d)
4.83 ± 0.17c (n: 47)
5.35 ± 0.18b (n:40)
5.55 ± 0.12a (n:60)
4.08 ± 0.17d (n:40)
5th instar (d)
5.61 ± 0.18c (n: 41)
5.59 ± 0.25c (n:40)
8.13 ± 0.37a (n:60)
7.20 ± 0.25b (n:40)
Male adult longevity (d)
51.55 ± 1.09c (n: 29)
48.54 ± 1.14d (n:19)
64.92 ± 2.02a (n:36)
62.76 ± 7.31b (n:19)
Female adult longevity (d)
85.25 ± 6.89d (n: 12)
100.29 ± 2.27c (n:21)
116.71 ± 2.34a (n:24)
109.21 ± 7.76b (n:21)
TPOP (d)
30.75 ± 0.64d (n: 12)
36.48 ± 0.64c (n:21)
39.62 ± 0.78a (n:24)
37.79 ± 0.20b (n:21)
APOP (d)
4.00 ± 0.25c (n: 12)
4.71 ± 0.53b (n:21)
4.46 ± 0.22b (n:24)
5.95 ± 0.50a (n:21)
Oviposition (d)
4.75 ± 1.18c (n: 12)
5.90 ± 0.58b (n:21)
6.88 ± 0.44a (n:24)
5.68 ± 0.22b (n:21)
Fecundity Egg/Female
126.50 ± 9.58d (n: 12)
178.64 ± 17.90b (n:21)
215.85 ± 9.27a (n:24)
162.95 ± 6.27c (n:21)
3.2 Population parameters
Longer mean relative growth rate was calculated on seeds Bt-Cyto-179 (P < 0.01). Significantly highest mean intrinsic growth rate was found on Bt-Cyto-179 as compared to other three tested seeds (P < 0.001) (Table 2). The life table parameters (Table 2) showed maximum values on Bt-Cyto-179 as compared to other three cotton seeds (P < 0.0001). The double time of O. hyalinipennis was significantly longer on CIM 620 (Table 1). Curves of sxj for O. hyalinipennis development on Bt and non-Bt cotton seeds were different (Fig. 1). Egg laying of O. hyalinipennis at x and j stages were different (Fig. 2). The Lx and Mx of O. hyalinipennis were greatly influenced by Bt and non-Bt cotton seeds (Fig. 2). The Ex of O. hyalinipennis longer on Bt-Cyto-179 seeds (Fig. 3). The Vxj of O. hyalinipennis adults have shown in the Fig. 4. r; The intrinsic rate of increase (per days). λ; The finite rate of increase (per days). GRR; Gross reproductive rate (offspring). R0; The net reproductive rate (offspring/individual). T; The mean generation time (days).
Parameters
Different cotton seeds
CIM 620
CIM 717
CIM 632
Bt-Cyto-179
r (per day)
0.06
0.07
0.07
0.07
λ (per day)
1.06
1.07
1.07
1.08
GRR (offspring)
123.86
154.94
89.23
155.35
R0 (offspring/individual)
15.18
37.52
30.96
51.80
Mean Generation Time (T) (Days)
44.49
51.08
49.59
56.39
Doubling Time (D) (Days)
11.34
9.77
10.01
9.90
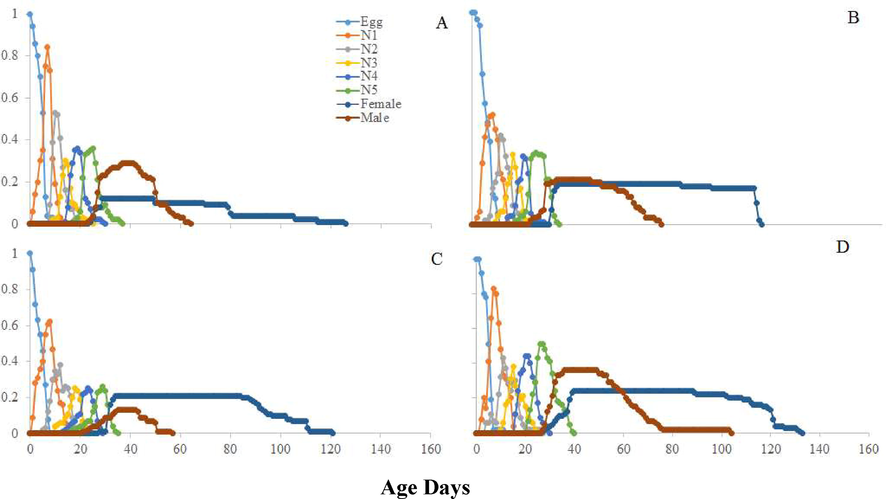
Influence of Bt and non-Bt seeds on the age-stage-specific survival rate (sxj) of the O. hyalinipennis. A: CIM 620, B: CIM 632, C: CIM 717, and D: Bt-Cyto-179, NI: 1st Instar Nymph, N2: 2nd Instar Nymph, N3: 3rd Instar Nymph, N4: 4th Instar Nymph, N5: 5th Instar Nymph.
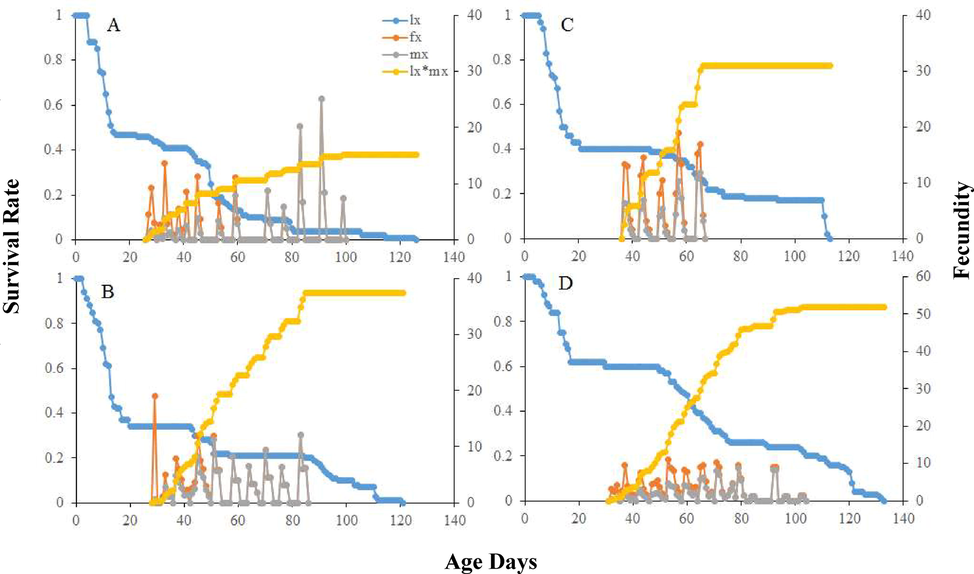
Influence of Bt and non-Bt seeds on the Age-specific survival rate (lx), female age-specific fecundity (fx), age-specific fecundity (mx), and age-specific maternity (lx*mx) of the O. hyalinipennis A: CIM 620, B: CIM 632, C: CIM 717, and D: Bt-Cyto-179.
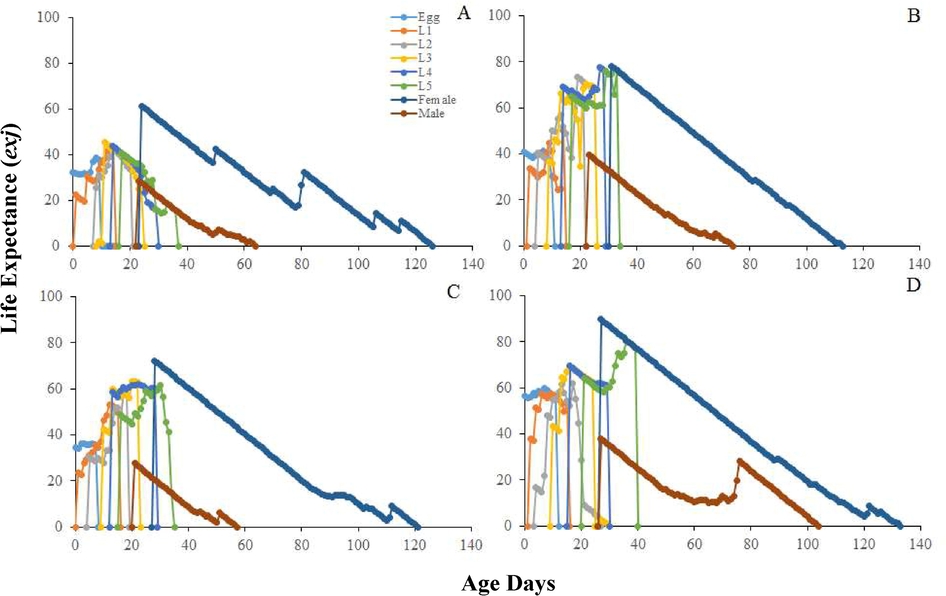
Influence of Bt and non-Bt seeds on the age-stage-specific life expectancy (exj) of the O. hyalinipennis A: CIM 620, B: CIM 632, C: CIM 717, and D: Bt-Cyto-179, NI: 1st Instar Nymph, N2: 2nd Instar Nymph, N3: 3rd Instar Nymph, N4: 4th Instar Nymph, N5: 5th Instar Nymph.
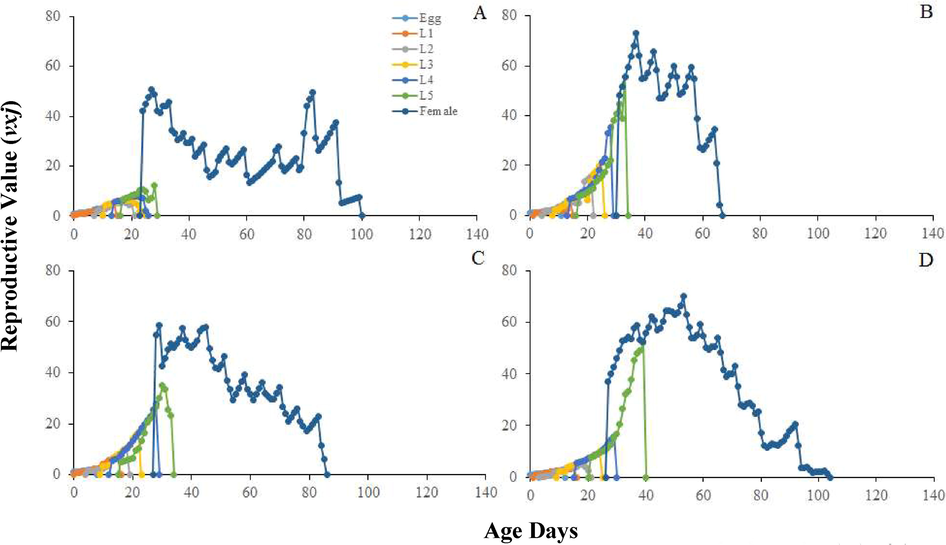
Influence of Bt and non-Bt different temperatures on the age-stage reproductive value (vxj) of the the O. hyalinipennis. A: CIM 620, B: CIM 632, C: CIM 717, and D: Bt-Cyto-179, NI: 1st Instar Nymph, N2: 2nd Instar Nymph, N3: 3rd Instar Nymph, N4: 4th Instar Nymph, N5: 5th Instar Nymph.
3.3 Protein contents
The protein contents were maximum in Bt-Cyto-179 (P < 0.0001) as compared to other three seeds (Fig. 5.).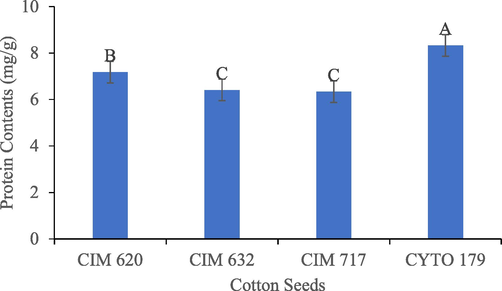
Protein Contents in Bt and non-Bt of cotton. Different letter bar represents the different values in the bars.
4 Discussion
Several studies explained the host influence on the development, demographic outcomes, and fitness of insects (Jaleel et al., 2018). This study was carried out by the considering the importance of host preference and the response of O. hyalinipennis. The difference in fitness and population parameters explains the preference of seeds. Growth rate in O. hyalinipennis enhanced on favorable hosts due to sufficient proteins in activation energy. As lot of studies revealed that Bt varieties of cotton are not resistant against sucking pests which resulted in enhanced metabolic rate, similar results were found in our study as Bt-Cyto-179 was more susceptible for dusky cotton bug.
Female adults kept specific behaviour of host selection for oviposition and development of newborns (Jaleel et al., 2018; Valladares et al., 2020). The olfaction and gustation sense of insect pests play an important role in their host selection and fitness of immature (Anderson and Anton, 2014; Jaleel et al., 2021a; Jaleel et al., 2021b). Maximum number of eggs was achieved on favorable seeds e.g., Bt-Cyto-179, this is possibly due to available seed contents. The adult fitness and fecundity are two main parameters for the increasing population of insects (Huang and Chi, 2012; Sarmad et al., 2020). A number of sucking pests undergo severe stress on unsuitable hosts.
The population parameters e.g., “r” can be better explained using two-sex programs (Huang and Chi, 2012; Jaleel et al., 2018). Fitness of insect pest explained the role of r, if its value becomes maximum than 0, than food of insect consider favorite or suitable for development and survival of insect (Chi et al., 2020). Another factor of increasing population is “R0” that explained the role of diet or food in high fecundity. In our study, the maximum eggs of O. hyalinipennis were recorded on Bt-Cyto-179 (Table 1). The results of this experiment summarized those seeds of Bt-Cyto-179 was found better for development of O. hyalinipennis so its life cycle becomes shorter with increase of fecundity. These results will be helpful in the management of O. hyalinipennis in Punjab, Pakistan. so further work would be carried out to find the molecular studies of O. hyalinipennis.
Acknowledgement
All authors’ acknowledgments to “Pakistan Central Cotton Committee” for its funding of this work by research project # “PSDP No.: 733” and entitled “Cotton Productivity Enhancement Through Eco-Friendly Pink Bollworm Management and Capacity Building PM Emergency Program” under “Ministry of National Food Security & Research, Islamabad (Through PSDP)”. The authors also extend their appreciation to the support of the Research Center for Advanced Materials Science (RCAMS) at King Khalid University, Abha, Kingdom of Saudi Arabia through a project number RCAMS/KKU/G003-21.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Climate Resilient Cotton Production System: A Case Study in Pakistan. Cotton Production and Uses. Springer; 2020. p. :447-484.
- Comparative growth rate, fecundity and behavioural diversity of the dusky cotton bug, Oxycarenus hyalinipennis Costa (Hemiptera: Lygaeidae) on certain malvaceous host plants. Proc. Indian Natl. Sci. Acad., B (Biol. Sci.). 1982;48:577-584.
- [Google Scholar]
- Experience-based modulation of behavioural responses to plant volatiles and other sensory cues in insect herbivores. Plant Cell Environ.. 2014;37(8):1826-1835.
- [Google Scholar]
- Impact of climate warming on cotton growth and yields in China and Pakistan: A regional perspective. Agriculture. 2021;11:97.
- [CrossRef] [Google Scholar]
- Characterization of genetic basis and realized heritability of bifenthrin-resistance selected in dusky cotton bug, Oxycarenus hyalinipennis (Costa)(Hemiptera: Lygaeidae) in Pakistan. Crop Prot.. 2021;141:105441
- [Google Scholar]
- Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol.. 1988;17:26-34.
- [Google Scholar]
- Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin.. 1985;24:225-240.
- [Google Scholar]
- Biological Control as a Key Tool for the Management of Invasive Species in Latin America and the Caribbean. Agricultural, Forestry and Bioindustry Biotechnology and Biodiscovery: Springer; 2020. p. :357-386.
- Biogenic synthesis of silver nanoparticles using propolis extract, their characterization, and biological activities. Sci. Adv. Mater.. 2019;11:876-883.
- [Google Scholar]
- Quality evaluation of Saudi honey harvested from the Asir province by using high-performance liquid chromatography (HPLC) Saudi J. Biol. Sci.. 2020;27(8):2097-2105.
- [Google Scholar]
- Hashmi I., Khan A. 2011. Adverse Health Effects of Pesticide Exposure in Agricultural and Industrial Workers of Developing Country. Pesticides—The Impacts of Pesticides Exposure. PP: 156.
- Age-stage, two-sex life tables of Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae) with a discussion on the problem of applying female age-specific life tables to insect populations. Insect Sci.. 2012;19:263-273.
- [Google Scholar]
- Realized heritability, cross-resistance and high risk of resistance development to spirotetramat in dusky cotton bug, Oxycarenus hyalinipennis Costa (Hemiptera: Lygaeidae), an emerging threat to BT cotton in Pakistan. Phytoparasitica 2021:1-11.
- [Google Scholar]
- Stability and fitness cost associated with spirotetramat resistance in Oxycarenus hyalinipennis Costa (Hemiptera: Lygaeidae) Pest Manag. Sci.. 2022;78(2):572-578.
- [Google Scholar]
- Optimization of treatment blocking the gustatory sense and feeding ethogram of red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae) to sugar. J. King Saud Univ. Sci.. 2021;33:101555
- [Google Scholar]
- Preference and effect of gustatory sense on sugar-feeding of fire ants. PeerJ. 2021;9:e11943
- [Google Scholar]
- Effects of temperature on baseline susceptibility and stability of insecticide resistance against Plutella xylostella (Lepidoptera: Plutellidae) in the absence of selection pressure. Saudi J. Biol. Sci.. 2020;27(1):1-5.
- [Google Scholar]
- Survey of Bt cotton in Punjab Pakistan related to the knowledge, perception and practices of farmers regarding insect pests. Int. J. Agric. Crop Sci.. 2014;7:10.
- [Google Scholar]
- Using two-sex life table traits to assess the fruit preference and fitness of Bactrocera dorsalis (Diptera: Tephritidae) J. Econ. Entomol.. 2018;111:2936-2945.
- [Google Scholar]
- Biodiversity and relative abundance of different honeybee species (Hymenoptera: Apidae) in Murree-Punjab, Pakistan. J. Entomol. Zool. Stud.. 2014;2:324-327.
- [Google Scholar]
- An investigation of the efficacy of hygienic behavior of various honey bee (Apis mellifera) races toward Varroa destructor (Acari: Varroidae) mite infestation. J. King Saud Univ. Sci.. 2021;33(3)
- [Google Scholar]
- Pollen source preferences and pollination efficacy of honey bee, Apis mellifera (Apidae: Hymenoptera) on Brassica napus crop. J. King Saud Univ. Sci.. 2021;33:101487
- [Google Scholar]
- Assessment of density-dependent feeding damage by the cotton dusky bug, Oxycarenus laetus Kirby (Hemiptera: Lygaeidae), in cotton. Turk. J. Agric. For.. 2014;38:198-206.
- [Google Scholar]
- Effect of constant and fluctuating temperature on the circadian foraging rhythm of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae) Saudi J. Biol. Sci.. 2021;28(1):64-72.
- [Google Scholar]
- Effects of elevated CO 2 on the fitness and potential population damage of Helicoverpa armigera based on two-sex life table. Sci. Rep.. 2017;7:1-13.
- [Google Scholar]
- Insect Pests of Cotton Crop and Management Under Climate Change Scenarios. Environment, Climate, Plant and Vegetation Growth. Springer; 2020. p. :367-396.
- Pesticide use and its impacts on human health and environment. Environ. Ecosyst. Sci.. 2020;4:47-51.
- [Google Scholar]
- Insect Pests of Cotton. Pests and their Management. Springer; 2018. p. :361-411.
- Razaq M., Mensah R., Athar Hu.R. 2019. Insect pest management in cotton. Cotton Prod. 85-107.
- Evaluation of the impact of genetically modified cotton after 20 years of cultivation in Mexico. Front. Bioeng. Biotechnol.. 2018;6
- [CrossRef] [Google Scholar]
- Toxicological studies on some important chemicals against Dysdercus koenigii Fabr (Hemiptera: Pyrrhocoridae) Pak. J. Zool.. 2016;48(5):1249-1254.
- [Google Scholar]
- Fitness and Predating potential of Antilochus coquebertii (Hemiptera: Pyrrhocoridae): A predator of the red cotton bug (Dysdercus koenigii) J. Kansas Entomol. Soc.. 2020;92:589-601.
- [Google Scholar]
- Stocks I., Peña J. 2013. Recent adventive scale insects (Hemiptera: Coccoidea) and whiteflies (Hemiptera: Aleyrodidae) in Florida and the Caribbean Region. Potential Invasive Pests of Agricultural Crops CAB International, Wallingford, United Kingdom: 342-362.
- Tuan S.J., Lee C., Chi H. 2014. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachishy pogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manage. Sci. 70: 805-813.
- Previous herbivory alerts conspecific gravid sawflies to avoid unsuitable host plants. Bull. Entomol. Res.. 2020;110(4):438-448.
- [Google Scholar]
- Inheritance mode and metabolic mechanism of the sulfoximine insecticide sulfoxaflor resistance in Oxycarenus hyalinipennis (Costa) Pest Manag. Sci.. 2021;77(5):2547-2556.
- [Google Scholar]




