

Translate this page into:
Timing of changes from a primitive reflex to a voluntary behavior in infancy as a potential predictor of socio-psychological and physical development during juvenile stages among common marmosets
*Corresponding author at: Department of Pediatrics, Saitama Medical University, 38, Morohongo, Moroyama-cho, Iruma-gun, Saitama, Japan. Tel.: +81 49 276 1418 koshiba@saitama-med.ac.jp (Mamiko Koshiba)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Available online 21 March 2015
Peer review under responsibility of King Saud University.
Abstract
With regard to socio-psychology, the complexity of a human’s neuronal function develops from initial primitive function to multimodal networking gradually through interaction between innate biological mechanisms and the surrounding environment. To study these developmental systems, we attempted to find a comparative-behavioral model among non-human primate species, common marmoset (Callithrix jacchus). In this research, we explored the correlation between the timing of change from involuntary- to voluntary-like movement in early stage and later the socio-emotional behavior, comparatively with the probable psychological development disabilities defined by our social behavior test (SBT). Using the ‘Infant Antigravity Task (IAT)’ and multivariate analysis, we quantitatively observed the neurological development from simple to complex behavior through interaction between motor and gravity environment. At the same time, we recorded the physiological and psychological development of two marmoset siblings (a male and a female) from their neonate stages to adult stages with their parents. In their lives, the male marmoset spontaneously showed his developmental delay in terms of body weight, blood glucose and socio-emotional difficulties under their own parental care. Based from the SBT results, the male demonstrated less social interaction with other family members compared to his sibling in the juvenile stage. Between these siblings, we looked for any predictable information in the earlier stage for future developmental issues particularly focusing on their behavioral expression during IAT from the 10th postnatal day until the 36th day.
Consequently, we found that both subjects expressed climbing-up behavior in the initial early period, but only the female who developed typically later, switched to jumping-down behavior with pre-facing to ‘down’ direction. Meanwhile, the male who would have developmental delay later, clearly did not show the switching pattern. The results suggest that the switch timing from involuntary to voluntary movement may be a possible predictor of juvenile and adolescent physiological and psychological retardation. The results also suggest that the primate model allows more methods to be developed for early detection of developmental disabilities that could be utilized in humans to pave the way for interventions and possible psychological or psychiatric treatment.
Keywords
Developmental milestones
Motivation
Gravity environment
Multivariate analysis
Body weight
Head surface temperature

- IAT
-
Infant Antigravity Task
- SBT
-
social behavior test
- bGlc
-
blood glucose concentration
- Thead
-
head surface temperature
- Theta 45
-
face preference to reference in SBT
Abbreviations
1 Introduction
The idea that earlier treatment during the development is the most powerful, has been agreed upon in the fields of education and medicine. Considering the development of our neuropsychological higher-order function, the biological basis may already start forming from the embryonic stage and it develops in infant, juvenile, adolescent and even adult and elderly stages step by step. These accumulated modules hierarchically integrated to form complex neurobiological information networks. In the earliest stage, humans express multiple kinds of primitive reflexes (Schott and Rossor, 2003; Blasco, 1994) as simpler central nervous systems of automatic regulation with sensory input from environment and motor output and interaction with environment. The primitive reflexes, including Moro reflex, grasping and rooting mostly appear transiently and disappear in the later developmental stages. Instead, more complex motor control behaviors appear, such as, ‘head control’, ‘rolling over’, ‘sitting’, ‘standing’ and ‘walking’ which are known as developmental milestones requiring higher-order nervous systems including voluntary function. In the series of motor developmental expression shifting, it has been reported possibility to predict the sign of psychological disabilities such as Asperger’s syndromes and autistic spectrum disorders by visualizing some behavioral characteristics and follow-up on the development (Teitelbaum et al., 2004; Jones and Klin, 2013). For innovation on non-established psychiatric treatment, we studied the neuronal development mechanisms from primitive to complex. The developmental process is thought as an interaction between biological and environmental mechanisms. We reported a possibility that the neonatal infant brain development affected by preterm metabolic conditions may predict the latter neuronal development of voluntary behavioral control (Koshiba et al., 2014) that is also relevant to the psychological development.
New world monkeys, common marmosets (Callithrix jacchus) are monogamous primates and make a small family for breeding. It is known that all family members cooperate with one another for breeding offsprings and we can often see the father taking care of the siblings except for feeding time during which the mother gives the offspring her own milk (Zahed et al., 2008). Their superior psychological capacity for socially cooperative communication (Flack, 2013) and altruistic behavior (Burkart et al., 2007) has led to the development of primate neurobiological systems in genetic and environmental interaction (Jokela et al., 2007) for studies of neurobiological psychiatry as an essential pre-clinical model (Okano et al., 2012) .
In this study, we firstly explain our evaluation of hypothetical diagnostic methods to quantitatively visualize the development of common marmosets focusing on their selection of antigravity relevant behavioral patterns that can be confirmed through the behaviors that appear or disappear over ages. A neonatal marmoset infant, after birth, grasps its parent’s body and clings to it continually to maintain the appropriate body temperature and to be protected from outside risks. The neonatal infants can reach the mother’s breast with its own control and parental supports under gravity loading. Their climbing ability may be considered as their primal capacity for grasping like humans do, by using the palmar and planter reflexes. A motor control and an activity test for the motor development in neonatal infants among common marmosets had been previously conducted with instrument and subjects’ climbing behaviors (Tardif et al., 2002). We furthermore challenged to discriminate the process from involuntary primitive to voluntary advanced function in their behaviors under a gravity environment. To seek any predictors in the behaviors, we selected the subjects who have been discriminated from their developmental troubles with our original quantitative analytic systems (Koshiba et al., 2011, 2013a,b,c,d,e; 2014, 2015; Senoo et al., 2011). The subjects of the study are two siblings raised by their own parents. In the results, we will firstly present the developmental history of the typical female and a male with spontaneous delay and describe our diagnosis, intervention and treatment results. Then, we will introduce our exploration on predictive factors in these siblings’ infant behaviors with our discrimination of three types, climbing-up, staying or jumping-down with quantification of their facing preference during Infant Antigravity Task (IAT). The results were analyzed using binominal regression analysis of generalized linear model (GLM) which could explain probabilities of explanatory parameters of their pre-facing and selecting behaviors in gravity.
2 Materials and methods
2.1 Animals
The experiment was approved by the Animal Care and Use Committee of Saitama Medical University. A pair of mature common marmosets (C. jacchus) whose previous siblings were removed from the family before their delivery, gave birth to triplets in October of 2013. One of the triplets died within 3 days. We tracked the development of the two other siblings (the male was named “Koma”, and the female was named “Yuzu”) as they were reared by their parents. The birth weights of Yuzu and Koma were 28.3 and 30.1 g, respectively. Provision of water, feeding balanced food pellet (CMS-1M, Clea, Japan), and home cleaning were performed from 10 to 11 a.m. daily. The infants were nursed until weaning was confirmed through their feeding behavior and body weight.
2.2 Repeated health and developmental checks
We tracked the health and developmental conditions of the two siblings by monitoring their body weight, blood glucose, and Infant Antigravity Task as described below. We also monitored their feces, daily activities, and body heat. To avoid separation anxiety or stress between siblings and parents, we completed a series of the measurements within 20 min between 3 and 4 p.m. The parents’ health conditions were similarly but less occasionally monitored.
2.2.1 Body weight
The subjects’ body weights were measured every day until postnatal day 10 (P10), and then every 2 days until P50, after which the subjects’ body weights were measured weekly for all subjects older than P50.
2.2.2 Blood glucose measurement
To record a representative information of developmental metabolic modulation every week after P51, we measured blood glucose utilizing ‘blood glucose meter’ (ARKRAY, Japan). We quickly sampled a small drop of blood from the caudal vein using a thin needle (27gage), while each infant subject was wrapped gently in our hands. The “sedation effect” (stress-less) of this procedure had been repeatedly experienced and confirmed among twelve marmoset infants (Koshiba et al., 2013e).
2.2.3 Infant Antigravity Task (IAT)
To investigate an infant marmoset’s physical development against gravity, we gave it Infant Antigravity Tasks (IAT) modified from the report of Clasping/Righting task developed by Tardif et al. (2002). Two types of cylinders made from aluminum cans were prepared by wrapping each circumferential surface with either a pee pad sheet for pets (SANOTEC, Japan; Sheet-cylinder) or a 4.4 mm-grid sheet (DAISO, Japan; Grid-cylinder).
As the initial condition of IAT, an experimenter let a subject cling to either of two cylinders and kept the long axis of the cylinder horizontally along and appropriately at a 200 mm height from a cushion ground to protect an infant from drops (Fig. 3A). The both sides of the cylinders were too smooth for the marmoset infants to climb, thus they could move up and down only at the sides of cylinders.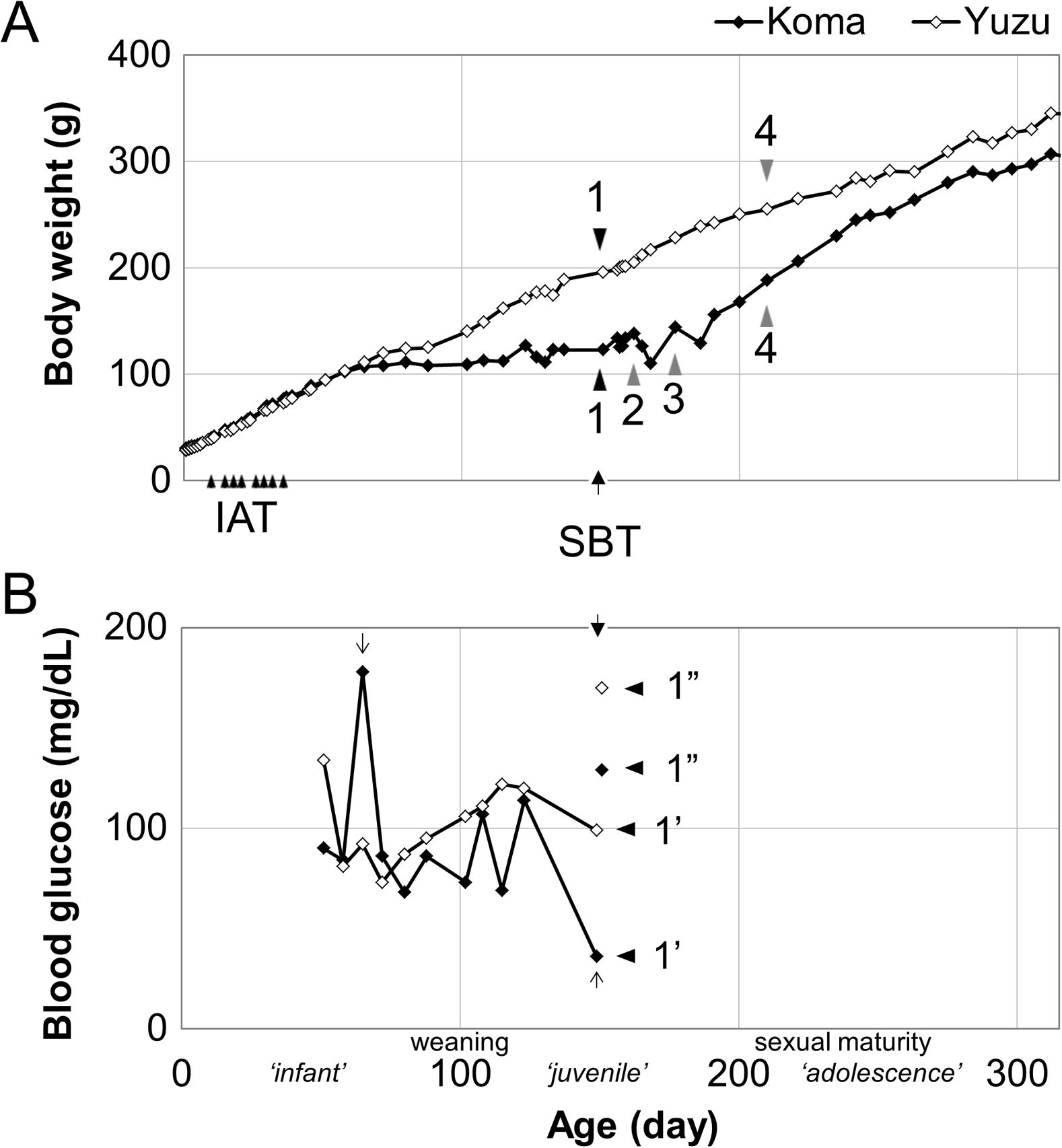
Age-dependent body weight and blood glucose concentration of siblings. (A) Body weight. All the events of this study are summarized as follows: IAT: Infant Antigravity Task (P10∼36). SBT: Social behavior test between a sibling and a parent at P150 when Yuzu started her independent life. 1: The worst condition of Koma. 2: Koma’s isolation. 3: Koma with parents again. 4: Both siblings together. (B) Blood glucose 1′: At the same time of one day before SBT 1″: Immediately following SBT. Fig. 2.
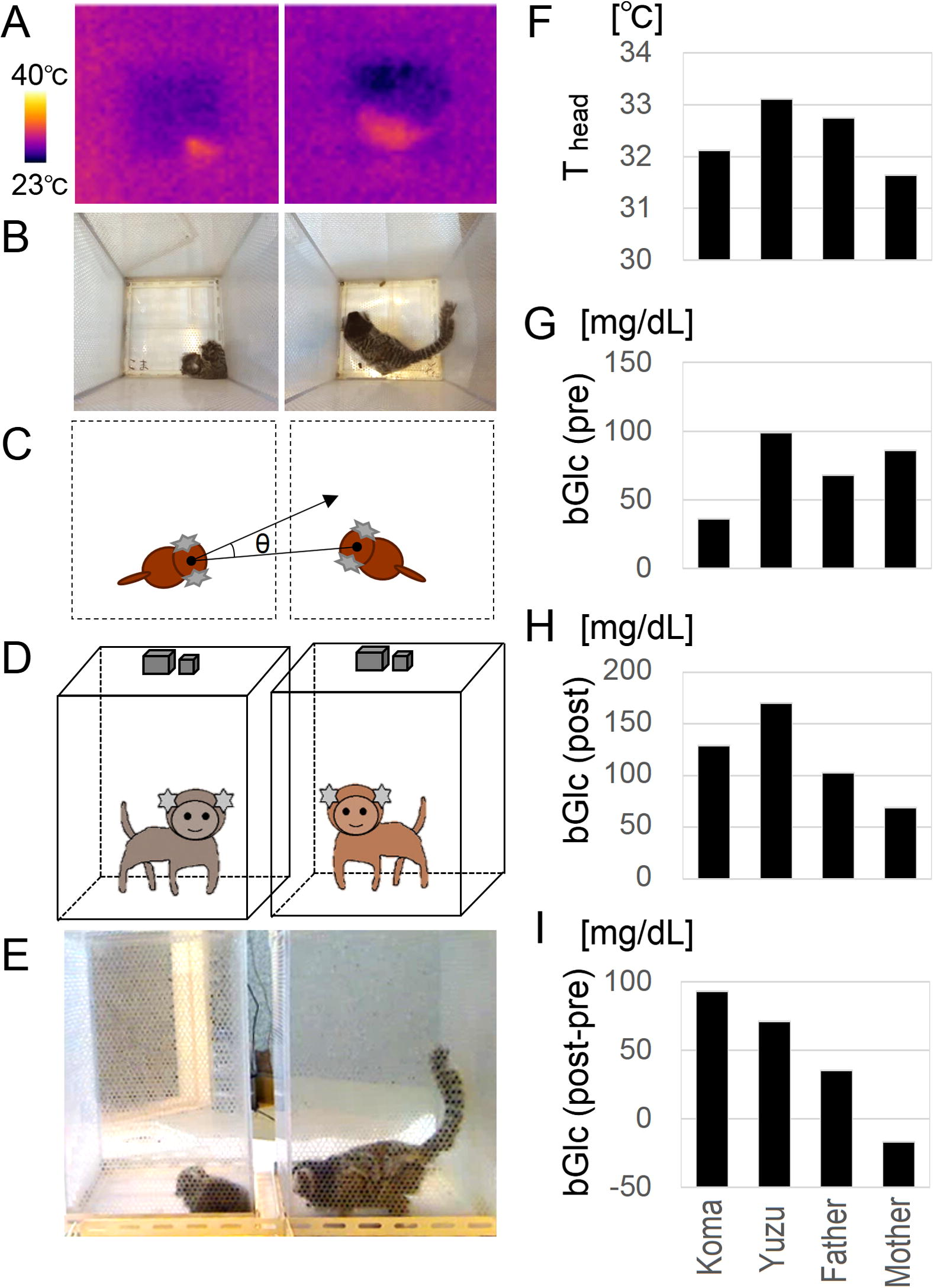
Social behavior test (SBT) among the family. (A) Thermo images acquired by the top simple sensors. (B) Images of web cameras at the top. (C) Definition of social face direction angle, theta. (D) Side views of C. (E) An image of a web camera at the sides. (F) Head surface temperature (Thead) during SBT. (G–I) Blood glucose concentration at one day before SBT (G), immediately after SBT (H) and their subtraction. Note the similar relative pattern of bar graphs between Thead (F) and bGl (post) (H), consistently.
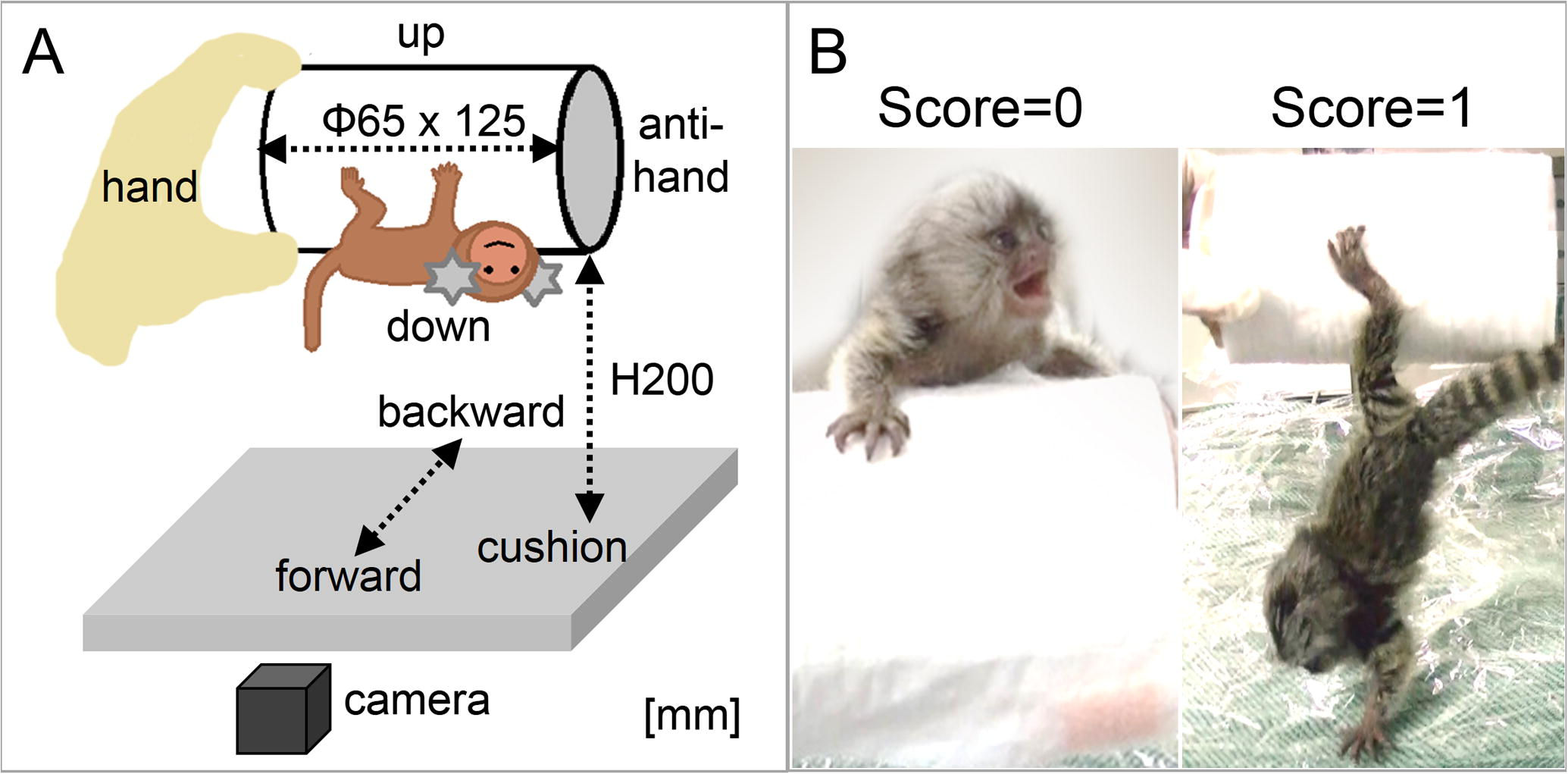
Infant Antigravity Task (IAT). (A) A set of IAT condition. A subject was set as this initial position at the beginning of each session. (B) Each example photo of either zero, “climbing-up (left)” or one, “jumping-down (right)”. Another case, staying on a cylinder within two-minute duration, was scored one.
An individual’s behavior for 2 min maximum was recorded by a digital video camera (SONY, Japan). We scored ‘one’ in either case that the marmoset actively jumped down to the cushion which is regarded as the end of session or kept staying on the cylinder, otherwise, we scored zero if the marmoset climbed using all its limbs to reach the upper part of the cylinder within the 2 min (Fig. 3B).
By using the video image (JPEG) per second acquired by an image software TMPGEnc VideoMasteringWorks5 (PEGASYS, Japan), we discriminated each individual’s face direction in six patterns of three dimensions (3D), “up” or “down”, “forward” or “backward” and “hand” or “anti-hand” (Fig. 3A), and summarized the time frequency of each direction during the test (Fig. 4).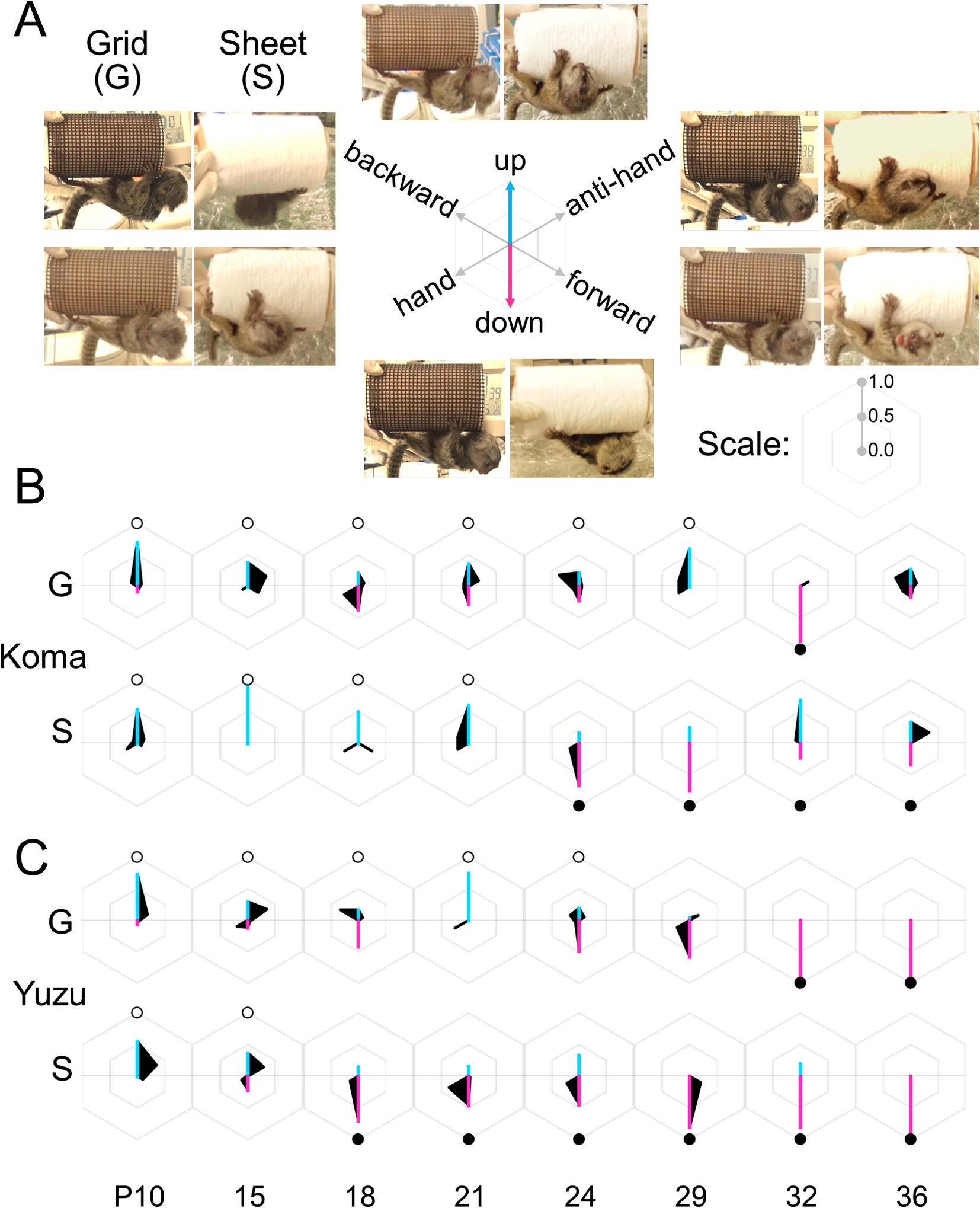
Age-dependent pattern shifts of face direction frequencies and the following behaviors, climbing-up, jumping-down or stay. (A) Each subject’s face preference was defined each of six directions (A, see also Fig. 3A,) in Infant Antigravity Task (IAT) with the Grid-cylinder (G) or Sheet-cylinder (S). (B and C) The hexagonal diagrams of Koma (B) and Yuzu (C) visualize their face preference pattern changes from postnatal day 10–36. An open circle at “up” in each diagram indicates a ‘climbing-up’ behavior, scored zero. A filled circle at “down” indicates ‘jumping-down’ to the cushion, scored one. No circle indicates ‘stay’ during a two-minute session, scored also one. Blue and pink lines in each graph emphasize face directions of “up” and “down”, respectively.
2.3 Social behavior test (SBT) Fig. 2
To generally check the state of each individual’s psychological and physical development at P150, we utilized the social behavior test (SBT) designed and evaluated by ourselves (Koshiba et al., 2008, 2011, 2013a,b,c,d,e, 2014, 2015; Senoo et al., 2011; Shirakawa et al., 2013). In a transparent cage made of perforated vinyl chloride walls (290 × 290 (bottom) × 790 (height) [mm]) with multiple 8 mm diameter holes for marmosets to climb and stay SBT was performed at between 9 and 11 a.m. The room temperature was set at 29 °C.
Each subject in a test cage met the mother, father or sibling one at a time in a random order in another cage for 50 s for visual, auditory and olfactory but not tactile interaction. A thermo camera (CHINO, Japan, Fig. 2A) on the top and visible light cameras (Logicool, Fig. 2B–E) on the top and sides of the cage recorded their behaviors. From the video images (JPEG) per second by using TMPEGenc (PEGASYS, Japan), the face direction of a subject in 3-D space coordinates was quantified using ImageJ (NIH, USA) and determined the relative face angle theta to a parent. If a subject faced a parent, the value of theta was zero [degree]. We defined a subject’s facing preference for each parent by the time ratio of taking the theta value less than 45° in the session (Fig. 2C, Koshiba et al., 2011, 2013a,b,c,d,e;Senoo et al., 2011; Shirakawa et al., 2013).
The head surface temperature of a subject was measured from the thermo image (Fig. 2A) every second and was corrected by the room radiant temperature and distance between the camera and the subject. The averaged value of a session was defined as the head surface temperature (Thead).
The blood glucose concentration (bGlc) of two siblings was measured by utilizing a ‘blood glucose meter’ (ARKRAY, Japan) a day before (pre) and immediately after (post) SBT.
2.4 Statistical analysis: generalized linear model (GLM)
To find the correlation of the jumping-down behavior during IAT with the face direction preference, we applied GLM assuming that the response variable climbing-up or jumping-down/staying on the cylinder follows binominal distribution. This is reasonable since the sum of the probability of the climbing-up and jumping-down/staying on the cylinder should be one and 0 given q as the probability of jumping-down/staying. We tested the two infant subjects (Names, Koma and Yuzu) on both cylinders per trial for 8 days over the developmental period from P10 until P36, accordingly 32 trials in total (N: the number of trial; N = 32) (Fig. 4). Given the jumping-down/staying count, one or zero in the trial independently as , it follows the binominal distribution below and obtains the likelihood function as the product of each probability where βj is regression coefficient defined below.
Using a binominal regression analysis, glm () provided by R (www.r-project.org), we can estimate based on maximum likelihood estimation and the best parameter selection by AIC (Akaike, 1974). In this estimation, we used logit-link function, , and the liner predictor described below,
In this formulation, (.) and denote explanatory variable and regression coefficient (weight of each explanatory variable) of each variable, respectively. We also estimated that the interaction between two variables was formulated as (Xi * Xj). Lastly, the categorical factor (name of subject) was introduced using a dummy function defined by “d = 1 given Name is Koma, and if Yuzu, d = 0”. We selected the best likelihood model by stepwise function using AIC (software R site).
3 Results
3.1 General diagnosis of Koma’s developmental delay in terms of body weight supposedly due to family problems and recovery with social intervention
The birth weights of Koma (male) and Yuzu (female) were 30.1 g and 28.3 g, respectively. Their body weight steadily increased until P65. The similarities in the development of their body weights were confirmed after the beginning several times of blood glucose measurement from P51 (Fig. 1A). At P65, Koma’s glucose value was extremely high, 180 [mg/dL] (Fig. 1B, a gray downward arrow), followed by continuous fluctuation. We could not identify any event speculated as the cause for Koma’s blood glucose increase. In parallel with this change, Koma’s body weight spontaneously began stagnation, on the while Yuzu’s weight kept increasing after the short plateau presumably because of weaning period and the blood glucose concentration stably increased until complete weaning. They lived with their parents in the same home cage until P150.
At P150, we decided to start our intervention to the family because Koma’s weight became almost half of Yuzu’s weight. Although we observed that both the siblings already begun eating pellets, Yuzu was observed clinging to her mother more often. Furthermore, we had repeatedly found Koma’s isolation from other members in the home cage. From these observations, we hypothesized the existence of social interaction problems such as competition between siblings for the mother’s breast milk even after the weaning period, or parental abuse or neglect. To diagnose their social conditions, we checked the socio-psychological states of each member of this family using our original diagnostic examination of social behaviors (SBT, Fig. 2) that we had designed and evaluated for a range of species, including the common marmoset. According to our previous studies (Koshiba et al., 2011, 2013a,b,c,d,e; Senoo et al., 2011; Shirakawa et al., 2013), we focused on their social face preference factor and head surface temperature which might possibly suggest subject’s emotional states in variable social contexts.
We computed the face preference score as described in Section 2 of this paper. Compared to Yuzu, who faced her parents equally, Koma faced his father less than his mother although the father faced Koma more but the mother faced either sibling equally (Table 1, “Infant to Parent” and “Parent to Infant”). More interestingly, regarding the infants’ facing other siblings, Yuzu never faced Koma at all, while Koma faced Yuzu very briefly (Table 1, “Infant to Infant”). The relative expression of head surface temperature (Fig. 2F) and blood glucose concentration after SBT (Fig. 2H, bGlc (post)) appeared similar among the family. The averaged head surface temperature (Thead) of Yuzu was higher than Koma’s and the parents’ (Fig. 2F), and blood glucose concentration was also the highest in Yuzu in both pre- and post-SBT (Fig. 2G and H). Meanwhile, the increase of blood glucose from pre- to post-SBT was the highest in Koma (Fig. 2I) which led our assumption to Koma’s psychological vulnerability at a novel context. A notable point was Koma’s strange behavior of crouching down that we had never seen in the previous SBT with a hundred of other common marmoset subjects. Because Koma’s body weight had been already lower, it was seriously considered as a life-threatening condition at that time. We generally decided to make that as the endpoint of any more diagnostic treatments including SBT and blood glucose concentration to avoid stress. According to the observed behaviors, we also adjusted the frequency of body weight and health checks to a minimum.
Category
Subject
Sex
Reference
Face preference (theta 45)
Infant to Parent
Koma
M
Father
0.67
Koma
M
Mother
0.80
Yuzu
F
Father
1.00
Yuzu
F
Mother
1.00
Infant to Infant
Koma
M
Yuzu
0.10
Yuzu
F
Koma
0.00
Parent to Infant
Father
M
Koma
0.30
Father
M
Yuzu
0.10
Mother
F
Koma
0.23
Mother
F
Yuzu
0.23
Referring to these general diagnosis of Koma and the other family members with socio-behavioral observation, we attempted to find the best social-environmental intervention for Koma’s recovery from developmental delay from P150, and it might imply the psychological background function of Koma’s developmental delay in terms of his body weight. Firstly, we only let Yuzu stay independently in a different cage because the age of the two siblings had already reached the age of independence from parental care (Fig. 1A, triangle 1). Consequently, only Koma could be taken care of by parents at an environment without another sibling which resulted in Koma’s weight increase. However, Koma also should be independent, so we attempted to let Koma become isolated from P162, with the expectation to relieve Koma from competition with Yuzu (Fig. 1A, triangle 2). This isolation environment, however, did not seem to fit Koma because of his weight loss. Therefore, we decided to let Koma stay with his parents again for a while from P177 (Fig. 1A, triangle 3) to give Koma the opportunity to gain weights before living with another sibling. It is confirmed that parental care leads to Koma’s developmental recovery in terms of weight gain. Eventually, both siblings were separated from the parents and lived together from P210 (Fig. 1A, triangle 4), leading to the typical weight development of both siblings with their independence from their parents. Currently, any members of this family are living well without any problems.
3.2 Seeking early predictors in behavioral development through Infant Antigravity Task (IAT)
We sought any hypothetically relevant factors in the siblings’ early behavioral records for predicting the future social problems from juvenile to adolescence. For this purpose, we focused on the neuronal psycho-cognitive development from primitive reactions to higher stage behavior such as voluntary movement and examined their behavioral expression at Infant Antigravity Task (IAT) from P10 to P36 as described in Section 2 of this study. The body weights of both siblings were equally increasing during this early period (Fig. 1A and Table 2).
Name
Cylinder
Age [day]
Up
Hand
Anti-hand
Backward
Forward
Down
Body weight [g]
Score
Koma
Sheet
10
0.537
0.195
0.122
0.073
0.073
0.000
40
0
Koma
Sheet
15
1.000
0.000
0.000
0.000
0.000
0.000
47.7
0
Koma
Sheet
18
0.500
0.250
0.000
0.000
0.250
0.000
49.4
0
Koma
Sheet
21
0.600
0.200
0.000
0.200
0.000
0.000
54.2
0
Koma
Sheet
24
0.167
0.167
0.000
0.000
0.000
0.667
58.7
1
Koma
Sheet
29
0.250
0.000
0.000
0.000
0.000
0.750
67.2
1
Koma
Sheet
32
0.682
0.000
0.000
0.091
0.000
0.227
71.6
1
Koma
Sheet
36
0.333
0.000
0.333
0.000
0.000
0.333
76.5
1
Koma
Grid
10
0.704
0.000
0.074
0.111
0.037
0.074
40
0
Koma
Grid
15
0.381
0.095
0.333
0.000
0.190
0.000
47.7
0
Koma
Grid
18
0.218
0.258
0.105
0.000
0.056
0.363
49.4
0
Koma
Grid
21
0.364
0.091
0.182
0.091
0.000
0.273
54.2
0
Koma
Grid
24
0.220
0.098
0.049
0.366
0.049
0.220
58.7
0
Koma
Grid
29
0.600
0.200
0.000
0.200
0.000
0.000
67.2
0
Koma
Grid
32
0.000
0.000
0.143
0.000
0.000
0.857
71.6
1
Koma
Grid
36
0.271
0.146
0.104
0.278
0.042
0.160
76.5
1
Yuzu
Sheet
10
0.550
0.000
0.350
0.000
0.100
0.000
39
0
Yuzu
Sheet
15
0.367
0.114
0.291
0.013
0.000
0.215
46
0
Yuzu
Sheet
18
0.150
0.150
0.000
0.000
0.000
0.700
48.5
1
Yuzu
Sheet
21
0.158
0.360
0.000
0.000
0.026
0.456
52.6
1
Yuzu
Sheet
24
0.333
0.222
0.000
0.000
0.000
0.444
56.9
1
Yuzu
Sheet
29
0.000
0.000
0.000
0.000
0.200
0.800
65.9
1
Yuzu
Sheet
32
0.200
0.000
0.000
0.000
0.000
0.800
68.9
1
Yuzu
Sheet
36
0.000
0.000
0.000
0.000
0.000
1.000
73.1
1
Yuzu
Grid
10
0.737
0.000
0.184
0.000
0.026
0.053
39
0
Yuzu
Grid
15
0.295
0.199
0.349
0.006
0.036
0.114
46
0
Yuzu
Grid
18
0.167
0.000
0.083
0.333
0.000
0.417
48.5
0
Yuzu
Grid
21
0.750
0.250
0.000
0.000
0.000
0.000
52.6
0
Yuzu
Grid
24
0.194
0.065
0.097
0.161
0.000
0.484
56.9
0
Yuzu
Grid
29
0.038
0.231
0.154
0.000
0.000
0.577
65.9
1
Yuzu
Grid
32
0.000
0.000
0.000
0.000
0.000
1.000
68.9
1
Yuzu
Grid
36
0.000
0.000
0.000
0.000
0.000
1.000
73.1
1
The coordination of visual direction and motor control is crucial to develop voluntary movement (Johnson, 1990). We analyzed the visual direction during IAT. The time frequencies of both siblings to six directions of three dimensions, “up” or “down”, “forward” or “backward” and “hand” or “anti-hand” (Fig. 3A) are represented in diagrams of Fig. 4. Generally, they preferred the face direction in “up”–“down” dimension than the others, “forward”–“backward” or “hand”–“anti-hand”. Both infants initially showed climbing-up behaviors (Fig. 4B and C, marked with white empty circles) but not jumping-down behaviors (Fig. 4B and C, marked black filled circles) at earlier ages. In the latter stages, both siblings were able to show jumping-down behaviors. The jumping-down behavior emerged earlier in the task with a Sheet-cylinder than a Grid-cylinder.
At the same time, we noticed that Yuzu shifted earlier from the climbing-up to the latter jumping-down behaviors than Koma. Koma’s behavior was not completely consistent with visual direction particularly at a sheet-cylinder. On and after P29 with a Sheet-cylinder or P32 with a Grid-cylinder, Yuzu showed jumping-down behavior while changing her face direction highly to “down” (Fig. 4C). On the other hand, Koma seemed to face not only “down” but also the other direction even at the cases of jumping-down with a Sheet-cylinder (Fig. 4B). Additionally, concerning the other two dimensions, Yuzu tended to face the “anti-hand” earlier, then, changed to “hand” direction later, however, Koma faced random directions. The data for this part of the study are shown in Table 2.
We finally evaluated whether there were any statistical differences among the face direction frequencies between Koma and Yuzu and if there could be an explanation to the significance of our score definition of jumping-down behavior, by generalized linear model (GLM) and stepwise selection of an optimal combination of explanatory variables (Table 3). Optimizing the coefficient of GLM formula, we obtained the following function. Score = −2.979−8.513 * Yuzu + 17.383 * down + −17.663 * hand + 45.496 * Yuzu * hand.
Estimate coefficient
Standard error
Probability
(Intercept)
−2.979
1.838
0.105
“Name(Yuzu)”
−8.513
4.634
(∗)0.066
“down”
17.383
7.510
∗0.021
“hand”
−17.663
11.914
0.138
“Name(Yuzu):hand”
45.496
26.874
(∗)0.090
We could confirm that the parameter of face direction frequency to “down” significantly correlated to the IAT score (the probability of jumping-down, p < 0.05). Furthermore, it was not significant but weakly showed the selection of “Name(Yuzu)” only but not Koma, suggesting different behavioral expressions between Koma and Yuzu in the explanation of jumping-down behaviors. The interaction between face direction frequency and “hand” shown as “Name(Yuzu) * hand” contributed weakly to jumping-down behavior (p < 0.1, Table 3), being consistent with the idea of the difference between two siblings.
4 Discussion
Our aim in this study was to explore any predictor of future problems in the psychological development and examine the correlation of the transition timing from primitive involuntary-like movement to voluntary-like movement in an early stage, with the trend of body weight recording that might be reflected in the psychological development in juvenile and adolescent states among common marmosets. While this species has been currently seen as a primate model for cellular and molecular biological studies (Sasaki et al., 2009; Marmoset Genome Sequencing and Analysis Consortium, 2014; Kilcoyne et al., 2014), these psychologically sensitive monkeys need to be aware of their human care-givers’ consideration for their life-long development with social and physical environments (Jaquish et al., 1997; Pryce et al., 2004; Dettling et al., 2002a; Senoo et al., 2011; Koshiba et al., 2011, 2013a, 2014; Shirakawa et al., 2013; Karino et al., 2015). In our marmoset model, we observed the spontaneous stagnation of body weight and fluctuation of basal blood glucose levels in a male juvenile marmoset beyond P65, while his female sibling developed well in these physiological parameters. The initial results of Fig. 1 about our social intervention trials and the success of recovery, like ‘loss and gain of function’, possibly suggest an approach using socio-environmental treatment and that the body weight trends might be generally useful to overview the developmental psychology concerning their vulnerabilities. Referring to the future problems concerning psychological vulnerability, we explored any sign in the infant’s physical responses against gravity.
To detect the differences in stress response from the earlier stages during postnatal day 10 through 36 in these subjects, we tried using the Infant Antigravity Task (IAT) to discriminate certain differences in behavioral patterns between the siblings using diagram visualization (Fig. 4). Although the disappearance of climbing-up behaviors after neonatal age had been reported in cotton-top tamarins and squirrel monkeys (King et al., 1974), essential factors and mechanisms of the behavioral and neurocognitive development have been unknown. Our result suggested that the developmental switching pattern from climbing-up at initial stage to jumping-down with pre-facing downward to the ground may be a predictor, since Yuzu showed this clear switch and had been typically independent from the parent beyond postnatal five months, on the while, Koma had developmental delay at the same age stage.
The age of behavioral switching from climbing-up to jumping-down was recorded earlier with a Sheet-cylinder than a Grid-cylinder. First of all, this switching may be related to the developmental change from righting and grasping reaction, the latter appeared even 24–36 h after birth (Tardif et al., 2002) to a more intentional behavior choice of jumping-down by suppressing the primitive reaction. The earlier switching with a sheet-cylinder than with a Grid-cylinder may be explained by the difference of the surface structure in terms of easiness of holding the body. However, the behavioral switching was observed with both cylinders.
In these tasks, we found the parameter of facing direction frequency to the ground “down” followed by jumping-down that was significantly explained by multivariate regression analysis, GLM. We regard that face direction almost means that their view direction in supposedly sensing the expected goal for deciding the following behavioral control. This series of processes must involve the subject’s voluntary and conscious behavior and motivation. On the other hand, climbing-up behavior was shown in a neonatal earlier stage of both siblings could not be correlated to their facing direction frequency. It is possibly one of instinctive primitive reflexes in common marmoset and this idea is supported in their neonatal nature of innate function in the initial 2 weeks to keep their warmth and safety within the parental body that is usually placed in the higher forest trees (Stevenson and Rylands, 1988). Because Yuzu showed jumping-down behavior from postnatal P18 with the smooth surface cylinder and P32 with the grid surface cylinder but not soon after birth, Yuzu might develop her motivation to the outer high-risk world, supposedly. It is consistent to visualize the common marmoset complex neuronal development in animal and environmental interaction considering gravity loading. It has been suggested that the primitive reflex in an earlier stage switched and disappeared among typical developing children and that their long persistence is prospective markers which may be a risk factor for Asperger’s syndrome (Teitelbaum et al., 2004). In our previous studies, some patterns in the behavioral development along with gravity direction may appear in human preterm infants, this is possibly a diagnostic marker of transition from primitive righting reflex to motivated behavior (Koshiba et al., 2013d).
An index of theta 45, social face preference at P150 was confirmed to estimate a social skill in common marmosets (Koshiba et al., 2011, 2013a). The factor, theta 45 was available for social diagnosis not only in marmoset but also among humans, especially to discriminate the behavioral pattern between those with Asperger’s syndrome and those who are typical subjects (Koshiba et al., 2013a). In this index, the mother’s social motivation to her infant was equal, but the father’s was higher to Koma rather than to Yuzu. On the other hand, while Yuzu always looked at both the mother and the father, Koma looked less at both parents and much less at the father. Since the developmental patterns of social skills possibly change depending on the interaction of parents and siblings (Koshiba et al., 2011, 2013a), it was possible to suppose that Koma experienced some psychological stress with his family at P51.
Concerning the subject siblings, the body weight transition is one of the gold standards for diagnosis of growth. Body mass index derived from body weight and height has been reported the correlation with cumulative adversity in childhood in cohort study (Slopen et al., 2014). We also confirmed the effectiveness of body weight as a biomarker of adult adaptability to climate and it seemed to reflect social experiences during the development (Karino et al., 2015). A previous study (Tardif and Bales, 2004) suggested that the birth weight positively and negatively correlated with the adult weight in twin and triplet, respectively. In our case, siblings were born as triplet and Yuzu having smaller birth weight (28.3 g) gained more weight at P300 than Koma having larger birth weight (30.1 g). This notion may not be the case; however, since the third sibling died at postnatal day 3 and the survived siblings grew as “twins” in reality. Considering the cause of developmental delay of body weight, it was reported that the rate of increase of body weight declines because of repeated parental deprivation until 4 weeks of age (Dettling et al., 2002b). In their study, the infant marmosets experienced repeated separation from their parents for a total of 9 h. They showed some stress responses such as an increase in cortisol, which was irrespective of their sex. In our study, the subjects experienced the separation within 20 min once every 3–5 days during Infant Antigravity Task (IAT) (P10–P36), so they experienced about 2 h and 40 min in total. Moreover, the developmental delay of body weight occurred only in the male infant, Koma and much later at P65 based on the IAT (Fig. 1A). Therefore, the effects of parental deprivation may not be a serious concern in our study. Law et al. (2009) stated that sensitivity to social stress is different between sexes. They suggested that, repeated parental deprivation led to the number of serotonin 1A (5-HT1A) receptor mRNA expression in the hippocampus which tended to decrease in females but increase in males. Moreover, social play negatively correlated with the expression of 5-HT1A receptor. In this study, since Koma’s social behavior to his parents was poorer than Yuzu’s (Table 1), the separation during IAT may have a little influence on Koma’s growth.
Tardif and Bales (2004) reported the case of the quite similar retardation of body weight gain from 2 months in their marmoset study, suggesting that the weaning-dependent change of weight gain related to the adult body weight. In our case, Yuzu and Koma showed similar sluggish weight gain around P60–P80 supposedly the beginning of the weaning period. Yuzu recovered her weight soon afterwards but Koma didnot. The exact reason of this body weight retardation is uncertain. Fluctuant blood glucose in Koma may be reflecting an irregular intake of the breast-milk because of socio-emotional problems between Koma and other family members. Physiologically, the capacity for urinary concentration develops in weaning rats irrespective of actual diet changes (Rane et al., 1985; Yasui et al., 1996). This result suggests that a systemic change may occur around a weaning period such as catabolism of carbohydrate and fatty acid metabolism. Therefore, it is possible that Koma may have some physiological vulnerability besides psychological stress.
Blood glucose concentration which is a biochemically measurable psychological stress marker (Armario et al., 1996), was measured every week from P51 to 123. Koma showed dramatic increase just before the trouble with his body weight began. This may suggest psychological stress in the family. While Yuzu’s blood glucose increased in parallel to her body weight, Koma showed fluctuating blood glucose levels and lower weight. This might have been the result of his deficiency in nutrient intake and his psychological developmental delay as expressed in SBT. Blood glucose also reflects the intake of food and the metabolic activity regulated in the endocrine systems such as the thyroid hormone, insulin, glucagon, and adrenal hormones (James and McFadden, 2004; Rafacho et al., 2014), which are required for growth and also psychologically affected through hypothalamic regulation and sympathetic nervous systems (Uresin et al., 2004; Sim et al., 2010). Koma might have had trouble because of feeding behaviors such as irregular intake of breast-milk and food pellets and the endocrine regulation during the weaning period.
We showed much earlier differences in switching age from primitive reflex against gravity to a kind of voluntary movement (jumping-down on the ground). The exact cause of this delay in the male neonate is uncertain. It is, however, quite reasonable to suppose that this delay of voluntary control function may cause a low chance to compete with sibling for feeding, more consuming energy otherwise passively supplied by the parent’s body heat, leading to higher anxiety and worsening of social interaction. Although we need a bigger sample size and detailed evaluation of the relationship among behavior, physiology and environment, it is possible that the difference of the physical development such as switching from climbing-up to jumping-down with pre-facing “down” in their earlier stages may be involved in the stress sensitivity or response. Our suggestions on this useful biomarker can help predict the socio-emotional development.
5 Conclusion
In the Infant Antigravity Task (IAT), both siblings expressed climbing-up behavior transiently at their neonatal age, however, only the female who would later develop typically switched to jumping-down with pre-facing-down behavior, but not the male who had developmental delay. These behavioral switching from primitive to voluntary motor control supported by generalized linear model (GLM) analysis may suggest a potential predictor of future physiological and psychological vulnerability.
Acknowledgements
We thank Dr. Aya Senoo and all the animal caregivers in our universities. We thank Prof. Hirohisa Kishino, University of Tokyo for leading our multivariate analysis. This work was partially supported by MEXT; JSPS KAKENHI Grant Number (21200017, 25119509, 25282221, JST-ALCA, JST-a-step) in Japan. We thank all members of our universities and institutes.
References
- A new look at the statistical model identification. IEEE Trans. Automic Control. 1974;AC-19:716-723.
- [Google Scholar]
- Acute stress markers in humans: response of plasma glucose, cortisol and prolactin to two examinations differing in the anxiety they provoke. Psychoneuroendocrinology. 1996;21(1):17-24.
- [Google Scholar]
- Primitive reflexes. Their contribution to the early detection of cerebral palsy. Clin. Pediatr. (Phila). 1994;33(7):388-397.
- [Google Scholar]
- Other-regarding preferences in a non-human primate: common marmosets provision food altruistically. Proc. Natl. Acad. Sci.. 2007;104(50):19762-19766.
- [Google Scholar]
- Early deprivation and behavioral and physiological responses to social separation/novelty in the marmoset. Pharmacol. Biochem. Behav.. 2002;73(1):259-269.
- [Google Scholar]
- Repeated parental deprivation in the infant common marmoset (Callithrix jacchus, Primates) and analysis of its effects on early development. Biol. Psychiatry. 2002;52(11):1037-1046.
- [Google Scholar]
- Understanding the processes behind the regulation of blood glucose. Nurs. Times. 2004;100(16):56-58.
- [Google Scholar]
- Interactions between infant growth and survival: evidence for selection on age-specific body weight in captive common marmosets (Callithrix jacchus) Am. J. Primatol.. 1997;42(4):269-280.
- [Google Scholar]
- Cortical maturation and the development of visual attention in early infancy. J. Cogn. Neurosci.. 1990;2(2):81-95.
- [Google Scholar]
- Serotonin receptor 2A gene and the influence of childhood maternal nurturance on adulthood depressive symptoms. Arch. Gen. Psychiatry. 2007;64(3):356-360.
- [Google Scholar]
- Attention to eyes is present but in decline in 2–6-month-old infants later diagnosed with autism. Nature. 2013;504:427-431.
- [Google Scholar]
- Common marmosets develop generation-specific peer social experiences that may affect their adult body weight adaptation to climate. Stress Brain Behav. 2015 in press
- [Google Scholar]
- Fetal programming of adult Leydig cell function by androgenic effects on stem/progenitor cells. Proc. Natl. Acad. Sci. U.S.A.. 2014;111(18):1924-1932.
- [Google Scholar]
- Development of early behaviors in neonatal squirrel monkeys and cotton-top tamarins. Dev. Psychobiol.. 1974;7:97-109.
- [Google Scholar]
- Development of an animal model and a social behavior assessment scoring method for developmental disorders. Ann. Gen. Psychiatry. 2008;7(Suppl. 1):S247.
- [Google Scholar]
- Reading marmoset behavior ‘semantics’ under particular social context by multi-parameters correlation analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2011;35:1499-1504.
- [Google Scholar]
- A cross-species socio-emotional behaviour development revealed by a multivariate analysis. Sci. Rep.. 2013;3:2630.
- [Google Scholar]
- Peer attachment formation by systemic redox regulation with social training after a sensitive period. Sci. Rep.. 2013;3:2503.
- [Google Scholar]
- Familiarity perception call elicited under restricted sensory cues in peer-social interactions of the domestic chick. Plos One. 2013;8(3):e58847.
- [Google Scholar]
- Socio-emotional development evaluated by Behaviour Output Analysis for Quantitative Emotional State Translation (BOUQUET): towards early diagnosis of individuals with developmental disorders. OA Autism. 2013;1(2):18.
- [Google Scholar]
- Blood glucose dependence on emotional behaviors and body surface temperatures in common marmoset’s socio-psychological learning with peers – for ‘Development of Human-Environment Interface by Sensing and Multivariate Analysis of Bio-Ecosystem’. ECS Trans.. 2013;50:9-14.
- [Google Scholar]
- Susceptible period of socio-emotional development affected by constant exposure to daylight. Neurosci. Res. 2014 S0168-0102(14)00222-3
- [Google Scholar]
- Early-infant diagnostic predictors of the neuro-behavioral development after neonatal care. Behav. Brain Res.. 2015;276:143-150.
- [Google Scholar]
- Early parental deprivation in the marmoset monkey produces long-term changes in hippocampal expression of genes involved in synaptic plasticity and implicated in mood disorder. Neuropsychopharmacology. 2009;34(6):1381-1394.
- [Google Scholar]
- The common marmoset genome provides insight into primate biology and evolution. Nat. Genet.. 2014;46(8):850-857.
- [Google Scholar]
- The common marmoset as a novel animal model system for biomedical and neuroscience research applications. Semin. Fetal Neonatal Med.. 2012;17(6):336-340.
- [Google Scholar]
- Deprivation of parenting disrupts development of homeostatic and reward systems in marmoset monkey offspring. Biol. Psychiatry. 2004;56(2):72-79.
- [Google Scholar]
- Glucocorticoid treatment and endocrine pancreas function: implications for glucose homeostasis, insulin resistance and diabetes. J. Endocrinol.. 2014;223(3):49-62.
- [Google Scholar]
- Development of urinary concentrating capacity in weaning rats. Pediatr. Res.. 1985;19(5):472-475.
- [Google Scholar]
- Generation of transgenic non-human primates with germline transmission. Nature. 2009;459(7246):523-527.
- [Google Scholar]
- The grasp and other primitive reflexes. J. Neurol. Neurosurg. Psychiatry. 2003;74(5):558-560.
- [Google Scholar]
- Effects of constant daylight exposure during early development on marmoset psychosocial behavior. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2011;35:1493-1498.
- [Google Scholar]
- Multivariate correlation analysis suggested high ubiquinol and low ubiquinone in plasma promoted primate’s social motivation and IR detected lower body temperature. J. Clin. Toxicol.. 2013;3:2.
- [Google Scholar]
- The regulation of blood glucose level in physical and emotional stress models: possible involvement of adrenergic and glucocorticoid systems. Arch. Pharm. Res.. 2010;33(10):1679-1683.
- [Google Scholar]
- Cumulative adversity in childhood and emergent risk factors for long-term health. J. Pediatr.. 2014;164:631-638.
- [Google Scholar]
- Neonatal behavioral scoring of common marmosets (Callithrix jacchus): relation to physical condition and survival. J. Med. Primatol.. 2002;31(3):147-151.
- [Google Scholar]
- Relations among birth condition, maternal condition, and postnatal growth in captive common marmoset monkeys (Callithrix jacchus) Am. J. Primatol.. 2004;62(2):83-94.
- [Google Scholar]
- Eshkol-Wachman movement notation in diagnosis: the early detection of Asperger’s syndrome. Proc. Natl. Acad. Sci.. 2004;101(32):11909-11914.
- [Google Scholar]
- Losartan may prevent the elevation of plasma glucose, corticosterone and catecholamine levels induced by chronic stress. J. Renin Angiotensin Aldosterone Syst.. 2004;5:93-96.
- [Google Scholar]
- Development of urinary concentrating capacity: role of aquaporin-2. Am. J. Physiol.. 1996;271(2 Pt. 2):461-468.
- [Google Scholar]
- Male parenting and response to infant stimuli in the common marmoset (Callithrix jacchus) Am. J. Primatol.. 2008;70(1):84-92.
- [Google Scholar]




