

Translate this page into:
Source, optimal dose concentration and longevity of trail pheromone in two Monomorium ants (Formicidae: Hymenoptera)
*Corresponding author. Address: Zoology Department, College of Science, King Saud University, P.O. Box 2455, Riyadh 11451, Saudi Arabia mmashely@ksu.edu.sa (Ashraf Mohamed Ali Mashaly)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Workers of Monomorium lepineyi and Monomorium bicolor secrete trail pheromones from their venom glands. The Dufour's gland and the hind gut secretions have no effect on the trail pheromone activity in M. lepineyi, while in M. bicolor the secretions have an inhibitory effect. The optimal dose of the trail- following in each of the mentioned two species was found to be 1.0 and 0.1 poison gland equivalent/30 cm trail, respectively. Although the concentration of the optimal dose in the two species is different, the longevity of each is nearly the same as the pheromone lasts 120 min.
Keywords
Ants
Monomorium lepineyi
Monomorium bicolor
Trail pheromone
Optimal dose
Longevity

1 Introduction
Species of the genus Monomorium are very diverse in size and habits, ranging from very small generalist scavengers to large, polymorphic seed harvesters. The genus is one of the most diverse ant genera of the family Formicidae, with more than 300 described species (Bolton, 1995; Heterick, 2001).
Diversity of exocrine glands are specialized for the production of chemical signals and are associated with particular cuticular structures which permit the release and dispersion of the secretion (Blum, 1985; Billen and Morgan, 1998). At least 39 different exocrine glands have been described in ants so far (Hölldobler and Wilson, 1990; Billen, 1994). Several of these represent a ‘standard exocrine set’, while others are thought to have only a limited distribution. In almost all species studied, trail pheromones originate from organs located in the gaster of the ants. The use of multiple trail pheromones by a single ant species means that foraging communication can be more complex than is possible with a single pheromone (Jackson and Ratnieks, 2006). Sources of trail pheromones are the venom gland, Dufour's gland and the hind tibia in subfamily Myrmicinae, pygidial gland in subfamily Ponerinae, Pavan's gland in subfamily Dolichoderinae, postpygidial gland in subfamily Aenictinae, and the hind gut in subfamily Formicinae (Billen and Morgan, 1998).
The present study aimed to investigate the source, optimal dose concentration and longevity of the trail pheromone in two Monomorium ants, Monomorium lepineyi and Monomorium bicolor.
2 Materials and methods
Monomorium ants comprised of few queens with brood, workers and males were collected from their natural raiding column (moisturized soil in a field) and immediately brought into the laboratory for trail-following experiments. The available species, M. lepineyi and M. bicolor were collected from a place 3 km north to Minia University, EL-Minia, Egypt.
Each ant species was kept in a plastic bottle containing moisturized soil. The bottle was placed in a plastic bowl with 25 cm internal diameter and 60 cm vertical wall to serve as a foraging area. Ants were fed with tiny droplets of bee's honey placed on a piece of paper and offered fresh dead insects at least once a week. Glass test tube filled with tap water and sealed with cotton piece was placed in the foraging area for ant drinking. To compensate moisture loss of the soil granules, few drops of water were added on alternate day.
The experimental determination of the glandular origin of trail pheromone involved presentation of extracts of the glands to ants. The behavioral response of 30 ants has been quantified through a circular trail test where the distance followed is equivalent to trail activity according to Pasteels and Verhaeghe (1974). Consequently, each of the dissected hind gut, venom gland and Dufour's gland was crushed in 100 μl hexane using a glass tissue grinder and immediately transferred by a Hamilton micro-syringe into the reservoir of a standard-graph pen, then the solution applied uniformly along the circumference of a circle with a diameter of 10 cm drawn on a sheet of white photocopying paper. The circumference of the circle was divided into arcs of 1 cm each. Hexane was used in the same manner as a control. Then the sheet of paper containing the extract was introduced into the foraging area and the mean number of arcs run by ants was calculated and used as a measurement of activity.
To determine the optimal dose of the trail pheromone of each species, different concentrations of the source gland were prepared (0.01, 0.1, 1, 5, 10, 20 and 40 glands equivalent per 30 cm trail). Each concentration in 100 μl hexane was allowed for workers in the foraging area for 20 min and the mean number of arcs run by individuals was calculated as in the previous test.
For determining the longevity of the trail pheromone, the optimum dose obtained from the previous experiment of each ant species was applied for workers after different time period from its initial application (0, 15, 30, 45, 60, 75, 90, 105, 120 min) and the mean number of arcs run by individuals was calculated.
Analysis of variance using one-way ANOVA (DSL) test was used to analyse the obtained data.
3 Results
3.1 The source of the trail pheromone
Data presented in Fig. 1 indicate that the poison gland is the source of the trail pheromone in M. lepineyi and M. bicolor where the mean number of arcs run by ant individuals was 23.37 ± 1.77 and 40.77 ± 2.98, respectively, whereas the Dufour gland or the hind gut of the two species induced a little activity and nearly similar to that evoked by a control blank bioassay.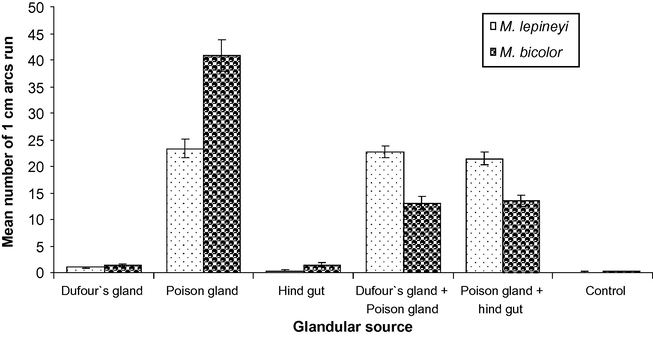
Trail-following activity evoked by different glandular sources of M. lepineyi and M. bicolor workers using the circular trail-following test.
In a trial to find out whether the Dufour's gland or the hind gut content has a synergetic effect for the venom secretion, a mixture of both the Dufour's and the poison gland or a mixture of both the hind gut and the poison gland was tested against workers. Results showed that the Dufour's gland and the hind gut secretions had no significant difference (P > 0.05 one-way ANOVA) in the trail pheromone activity in M. lepineyi while in M. bicolor the activity was significantly (P < 0.01 one-way ANOVA) decreased being less than that of poison gland extract alone (Fig. 1).
3.2 Effect of concentration on trail-following activity
Data presented in Fig. 2 indicate that workers of M. lepineyi and M. bicolor utilize a concentration of 1.0 and 0.1 gaster extract/30 cm trail, respectively, to evoke their highest activity. The activity was significantly decreased at a concentration below and above 1.0 and 0.1 gaster (P < 0.01 one-way ANOVA), respectively.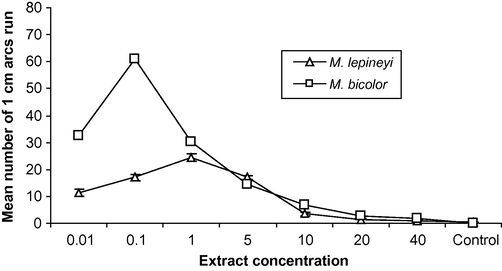
Response of M. lepineyi and M. bicolor workers to different concentrations of gaster extract.
3.3 The longevity of the pheromone
To determine how long the pheromone of M. lepineyi and M. bicolor can persist, the optimum dose obtained in the previous experiment was allowed for workers after different period of time after its initial application. It was found that the activity of workers toward the optimum dose of M. lepineyi (1.0 gaster) and M. bicolor (0.1 gaster) decreased gradually by the time until reached its lowest level after 2 h (Fig. 3).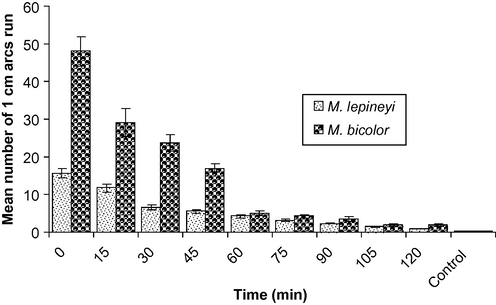
Longevity of the trail pheromone of the ant M. lepineyi and M. bicolor (1 and 0.1 gaster equivalent/30 cm trail, respectively).
4 Discussion
In this study the poison gland has been identified as a source of the trail pheromone in M. lepineyi and M. bicolor as in other Monomorium species, Monomorium floricola, Monomorium minimum Buckley and Monomorium pharaonis L. (Blum, 1966). Ritter et al. (1973) reported that M. pharaonis has been found to produce two trail substances from the poison gland; which were identified to be Monomorine I and Monomorine III. However, the true pheromone was identified as faranal secreted from the Dufour's gland and shown to be much more active than the monomorines (Ritter et al., 1977). When a mixture of both the poison gland and the Dufour's gland or hind gut was tested as a trail for M. lepineyi, no significant difference in activity of workers was obtained as in some Pheidole spp. (Ali and Mashaly, 1997a) whereas in M. bicolor workers, the activity was significantly decreased. In this case is probably due to the presence of certain substance (s) in the Dufour's gland or the hind gut which acts as an inhibitor as in Tetramorium simillimum (Ali and Mashaly, 1997b).
Ants are able to optimize their foraging behaviour by selecting the most rewarding source, due merely to a modulation of the quantity of pheromone laid on a trail (Hangartner, 1969; Hölldobler and Wilson, 1990; Traniello and Robson, 1995). Data in this study indicate that workers of M. lepineyi and M. bicolor utilize a concentration of 1.0 and 0.1 gaster equivalent per 30 cm, respectively, to evoke their highest activity. Van Vorhis Key et al. (1981) demonstrated that the optimal activity was found in response to a trail containing of 0.1–1.0 ant equivalent per 50 cm for Iridomyrmex humilis Mayer. In Tetramorium impurum Forester, the highest activity was reported at a concentration of 0.1 poison gland equivalent/30 cm trail (Morgan et al., 1990). The optimal dose of trail-following activity of Pheidole teneriffana which induced the highest activity was only one gaster equivalent/30 cm (Ali, 1996). Pheidole jordanica, Pheidole sinatica and Pheidole sp., utilize a wide range of pheromone concentration as the highest activity was fallen in between 1 and 5 gasters equivalent/30 cm trail (Ali and Mashaly, 1997a). Also the same range was observed with Crematogaster inermis Mayr, which utilizes the tibial gland as the source of the pheromone, while Leptothorax angulatus Mayr and T. simillimum Smith, utilize moderately higher concentration as the optimal dose was found to be five gasters for the same distance of trail (30 cm) (Ali and Mashaly, 1997b).
A parameter critical to the function of a trail is its persistence, and trail longevity must be matched to the foraging ecology of a particular species. Indeed, in ants trail longevity varies from minutes in Aphaenogaster albisetosus (Hölldobler et al., 1995) to several weeks in some Eciton species (Torgerson and Akre, 1970). Short-lived trails can rapidly modulate recruitment to ephemeral food sources, whereas long-lived trails will be more suited to persistent, or recurrent, food sources. At the optimal dose of the trail pheromone of M. lepineyi and M. bicolor, the activity of workers decreased to its lowest level after 2 h (Fig. 3).
The trail of M. pharaonis Linnaeus remained active for about a day, while those of M. minimium Buckle, were hardly active after 2.5 h (Blum, 1966). Ali (1996) found that, in Ph. teneriffana Forel, the optimal dose of the trail completely disappeared after 1 h. In Ph. jordanica, Ph. sinatica and Ph. sp., the activity of workers decreased to its lowest level after 75–90 min from its initial application at a concentration of two gasters, the activity recorded just a trace after 105–120 min at a concentration of five gasters (Ali and Mashaly, 1997a).
Acknowledgment
We are very grateful to Prof. Dr. C.A. Collingwood, (City museum, Leeds, UK) for identification of the ants species.
References
- Source, optimum dose response, longevity and isolation of trail pheromone of the ant Pheidole teneriffana (Forel) (Formicidae: Hymenoptera) J. Egypt. Ger. Soc. Zool.. 1996;20(E):69-82.
- [Google Scholar]
- Trail pheromone investigation of some Pheidole ants (Formicidae: Hymenoptera) Egypt. J. Zool.. 1997;28:113-123.
- [Google Scholar]
- Study on trail pheromone of three Myrmicine species, Leptothorax angulatus Mayr, Tetramorium simillimum Smith and Crematogaster inermis Mayr (Formicidae: Hymenoptera) J. Egypt. Ger. Soc. Zool.. 1997;24(E):1-16.
- [Google Scholar]
- Billen, J., 1994. Morphology of exocrine glands in social insects: an update 100 years after Ch. Janet. In: Lenoir, A., Arnold, G., Lepage, M. (Eds.), Les Insectes Sociaux. Publications Université Paris Nord, Paris, p. 214.
- Pheromone communication in social insects: sources and secretions. In: Vander Meer R.K., Breed M.D., Winston M.L., Espelie K., eds. Pheromone Communication in Social Insects: Ants, Wasps, Bees, and Termites. Boulder, Colo.: Westview Press; 1998. p. :3-33.
- [Google Scholar]
- The source and specificity of trail pheromone in Termitopone, Monomorium and Huberia and their relation to those of some other ants. Proc. R. Entomol. Soc. Lond. 1966:155-160.
- [Google Scholar]
- Exocrine systems. In: Blum M.S., ed. Fundamentals of Insect Physiology. New York: John Wiley; 1985.
- [Google Scholar]
- A New General Catalogue of the Ants of the World. Cambridge, Massachusetts: Harvard Uni. Press; 1995. p. 504
- Structure and variability of the individual odor trail in Solenopsis geminata Fabr. (Hymenoptera, Formicidae) Z. Vgl. Physiol.. 1969;62:111-120.
- [Google Scholar]
- Revision of the Australian ants of the genus Monomorium (Hymenoptera: Formicidae) Invertebr. Taxon.. 2001;15:353-459.
- [Google Scholar]
- The Ants. Berlin, Heidelberg, London, Paris, Tokyo, Hong Kong: Springer; 1990. p. 269
- Recruitment pheromones in the ants Aphaenogaster albisetosus and A. cockerelli (Formicidae: Hymenoptera) J. Insect Physiol.. 1995;41(9):739-744.
- [Google Scholar]
- Trail pheromone of the ant Tetramorium impurum and model compounds: structure–activity comparisons. J. Chem. Ecol.. 1990;16(12):3493-3510.
- [Google Scholar]
- Dosage biologique de la pheromone de piste chez les fourageuses et les reines de Myrmica rubra. Ins. Soc.. 1974;21:167-180.
- [Google Scholar]
- 5-Methyl-3-butyl-octahydroindolizidine, a novel type of pheromone attractive to Pharaoh's ants (Monomorium pharaonis (L.)) Experientia. 1973;29:530-531.
- [Google Scholar]
- Trail pheromone of the Pharaoh's ant, Monomorium pharaonis: isolation and identification of faranal, a terpenoid related to juvenile hormone II. Tetrahedron Lett.. 1977;30:2617-2618.
- [Google Scholar]
- The persistence of army ant chemical trails and their significance for the Ecitonine ecitophile association (Formicidae: Ecitonine) Melanderia. 1970;5:1-28.
- [Google Scholar]
- Trail and territorial pheromones in the social insects. In: Bell W.J., Cardé R., eds. The Chemical Ecology of Insects. Vol vol. II. Berlin: Chapman & Hall; 1995. p. :241-285. (Chapter 7)
- [Google Scholar]
- Effect of gaster extract trail concentration on the trail following behavior of the Argentine ant Iridomyrmex humilis (Mayr) J. Insect Physiol.. 1981;27:363-370.
- [Google Scholar]




