

Translate this page into:
Optimizing diet thickness and egg density for economic mass rearing of Ephestia kuehniella Zeller, 1879 (Lepidoptera: Pyralidae): A laboratory host for biological control agents
⁎Corresponding author. mehmetmamay@hotmail.com (Mehmet Mamay)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Background
Mediterranean flour moth [Ephestia kuehniella Zeller, 1879 (Lepidoptera: Pyralidae)] (MFM hereafter) is a significant pest of stored products, particularly flour and causes significant economic damages. However, its eggs are important laboratory host for mass rearing of biological control agents of important agricultural pests. However, diet thickness and eggs’ density to be used in a specific area for economic mass rearing are not fully understood.
Methods
This study optimized the diet thickness and eggs’ density to be used in a specified area for harvesting higher number of MFM eggs with low cost. The MFM was reared under dark environment, i.e., 25 ± 1 °C temperature and 60–70% relative humidity in 32×26×6 cm plastic tubs. The diet consisted of wheat flour and wheat bran in 2:1 ratio by weight, respectively. Three diet thicknesses [i.e., 0.5 cm (thin), 1.5 cm (medium) and 2.5 cm (thick)] and three egg densities [i.e., 25 mg (low), 50 mg (medium) and 75 mg (high)] were tested to determine the most economic and productive combination.
Results
Life history traits, i.e., moths’ production, moths’ ratio and egg production were significantly altered by individual an interactive effect of diet thickness and egg density. The highest number of moths was obtained for medium diet thickness and high egg density. Medium diet thickness with low egg density resulted in the lowest number of moths. The highest number of eggs were produced by thin and medium diets and high egg density combinations, and these combinations proved the most economic for mass production of 100 g eggs.
Conclusion
The results of the current study indicated that combination of medium and thin diet thicknesses with high egg density are the most economic for mass production of MFM. Interestingly, the study revealed that terminating moths’ collection at 2nd week after first moth emergence and egg collection at 4 days after the moths are placed in egg laying containers is the most suitable time for the economic mass rearing of MFM. However, egg densities >75 mg need further testing on thin and medium diet thicknesses to explore whether they are more economic than recommended densities in this study.
Keywords
Ephestia kuehniella
Mass rearing
Biological control
Laboratory host
Egg yield

1 Introduction
Rapid population growth, increased urbanization, and decrease in agricultural areas have been decreasing agricultural products per capita. Poor pest control are the other reason of decreasing agricultural productivity (Wu et al., 2011). Weeds are the most damaging pests globally causing 14.7% losses in agricultural production annually followed by insect pests (11.2%) and diseases (9.1%) (Oerke, 2006). These losses are equivalent to one-third of global agricultural production. If 6–12% post-harvest losses are added, total losses reach 40–48% (Dhaliwal et al., 2015). Several methods are used to manage the losses caused by these pests depending on production conditions, technology, and economic conditions of the producers (Upholt, 1980).
Chemical control with pesticides is the most quick and effective method of pest management opted by the farmers (Cherif et al., 2021). However, it poses significant negative effects on environment and human health (Sönmez and Mamay, 2018; Rani et al. 2021). Frequent use of chemicals results in pesticide residues (Mehmood et al., 2021; Philippe et al., 2021), results in the evolution of pesticide resistance in various insect species (Mamay and Mutlu 2019; Arthur et al. 2021; Ijaz and Shad 2021), disturbs natural balance through killing non-target natural enemies (Targanski et al., 2021), causes phytotoxicity in crop plants, increases management costs, and causes air (Guida et al., 2021), soil (Alshemmari et al., 2021) and water pollution (Senthil Kumar and Janet Joshiba, 2021). These reasons have forced the world to switch towards environment-friendly alternative pest management methods. Biological control is considered environment-friendly pest management method since decades (Bale et al., 2008; Barratt et al., 2018; Chandler et al., 2011; Mamay and Mutlu, 2019; Messing and Brodeur, 2018; van Lenteren et al., 2018). It deals with the management of insect pests, diseases, and weeds through natural enemies (Cherif et al., 2021). In short, it is the use of beneficial organisms against harmful pests (Bale et al., 2008). Biological control agents are grouped into three groups, i.e., predators (predator bugs), parasitoids (parasitic bugs) and pathogens (Rhodes, 1993).
The Mediterranean flour moth [Ephestia kuehniella Zeller, 1879 (Lepidoptera: Pyralidae)] is one of the most destructive cosmopolitan pests of stored grains, particularly flour (Kurtuluş et al., 2020; Özder, 2004; Xu et al., 2007). Mediterranean flour moth (MFM, hereafter) is distributed in all parts of the world, although endemic in the Mediterranean basin (Kurtuluş et al., 2020). It is regarded as an important pest since it is capable of damaging both flour and the whole grain of cereals (Nouri et al., 2019; Salehi et al., 2014). The webbings produced by MFM larvae of MFM obstruct mechanical operations. The consumers often reject cereal products having webbings and larval infestation of MFM (Hansen and Jensen, 2002).
Although MFM is an economically important stored products’ pest (Nouri et al., 2019), its eggs and larvae are frequently used as a conventional laboratory host for the rearing of many entomophagous insects (Morales-Ramos et al., 2014; Samara et al., 2008) such as parasitoids (Kurtuluş & Kornoşor 2015; Borzoui et al. 2016) and predators (Serkan et al. 2017; Abdalla et al. 2018; Sönmez and Mamay, 2018; Mamay and Mutlu, 2019). The requirement of mass rearing host or prey of biological control agents (natural enemies) doubles the rearing cost as both species are reared under laboratory conditions (Van Driesche and Bellows, 1996). The MFM is among the most preferred laboratory hosts for the scientific and commercial rearing of natural enemies because of its relatively short lifespan, high fecundity rate (200–300 eggs per female) and ability to grow on simple/economical diet. Although MFM can be reared on cheap diet, diet has a significant impact on the life history traits of insect species (Awmack and Leather, 2002; Umbanhowar and Hastings, 2002). The survival and productivity of insect species is significantly altered by the diet quality (Awmack and Leather, 2002). Therefore, inferring the impact of diet quality and quantity on the life history traits could provide valuable insights and helps in the rapid mass rearing of the target species. Several earlier studies have reported that different diets had significant impact on the life history traits of MFM (Faal-Mohammad-Ali and Shishehbor, 2013; Kurtuluş et al., 2020; Tarlack et al., 2015). However, rare or no studies have been conducted on the interactive effect of diet thickness and egg density on egg production of MFM. Nonetheless, no study has computed the costs incurred on the rearing of MFM.
Several studies have inferred life history traits of MFM on varying diets, none of these investigated the diet quantity and economics of the mass production. This study determined the impact of different diet thicknesses and egg density on egg production and costs of rearing. Finding the most economic diet thickness and egg density combination for economic rearing of MFM was the major objective of the study. It was hypothesized that rearing cost will increase with increasing diet thickness and egg density. The results of the study would help towards economic mass rearing of MFM, which serves as a laboratory host of biological control agents.
2 Materials and methods
2.1 Experimental site
The current study was conducted in the controlled insectarium located at Department of Plant Protection, Faculty of Agriculture, Harran University, Şanlıurfa Turkey.
2.2 Experimental material
The main materials used in the study were eggs and moths of MFM, and wheat flour and bran. The working materials included plastic tubs measuring 32×26×6 cm, white cheesecloth, and rubbers to cover the tubs, scales, egg trays covered with gauze, brushes, strainers, an oven and aspirator in which flour and bran mixture was sterilized.
2.3 Mass rearing of Ephestia kuehniella
The MFM was reared under 25 ± 1 °C temperature and 65 ± 5% relative humidity. Flour-bran mixture at 2:1 ratio was used as a culture medium (Bulut and Kılınçer, 1987). The flour-bran mixture was sterilized in an oven at 60 °C for 3–3.5 h and stored in a refrigerator at 4 °C. Approximately 2 kg of this ready mix was placed in plastic tubs (27×37×7 cm) where necessary. Approximately 50 mg of MFM eggs were sprinkled into each plastic tubs and the tubs were covered with white tulle. Adults emerging 35–40 days after the culture were collected with an aspirator and placed in plastic egg-laying containers with wire rims to lay eggs. These containers were laterally placed in plastic tubs having white paper at the bottom to easily collect the eggs laid by MFM. The eggs were collected from these containers with two days interval. Some of the freshly collected eggs were used for MFM culture, while the rest were stored in a deep freezer at −20 °C.
2.4 Impact of different diet thicknesses and egg densities on moth and egg production
The MFM rearing was done following the procedure of Bulut and Kılınçer (1987). Flour-bran mixture at 2:1 ratio was used as a culture medium/diet. Three different diet thicknesses, i.e., 0.5 (thin), 1.5 (medium) and 2.5 cm (thick) were used in the study. Furthermore, three different egg densities, i.e., 25 (low), 50 (medium) and 75 mg (high) were included in the experiment. Several studies focusing on mass rearing used 50 mg egg density; therefore, a higher and lower density was tested in this study (Mamay and Mutlu, 2019; Dusak, 2018). The experiment was conducted according to randomized complete block design with factorial arrangement. Diet thickness was taken as the main factor, whereas egg densities were regarded as sub factor. All treatments had 30 replications and experiment was repeated over time (two experimental runs for each treatment). The diet was sterilized and stored as described above. For different diet thickness treatments, the diet was spread in the tubs to the desired thickness according to the treatment and then eggs with different densities were sprinkled over diet to complete the rearing procedure. The 0–24 h-aged eggs obtained from the stock culture were cleaned with a soft brush and strainer. The desired quantity of eggs according to the treatments was weighed with a sensitive electronic balance and homogenously sprinkled on the diet. The tubs were then covered with cheesecloth which was fixed with the help of rubber. The experiment was observed every second day till 30 days. The number of moths completing their development were counted from each treatment combination. The number and days of first and last adult emergence were also recorded. Total number of moths emerged from each treatment as well as weekly average was computed and interpreted.
The emerging moths were collected with an aspirator and shifted to plastic egg laying containers as described above. The eggs laid after 3rd and 5th day of shifting were collected with the help of a delicate brush and cleaned by passing through a sieve. The harvested eggs from each treatment were weighed on an electronic balance.
The ratio of moths emerged at different weeks was computed to infer the optimum termination time for the experiment. The ratio of the moths emerged at 1st, 2nd, 3rd and 4th week to the total number of moths was computed.
2.5 Statistical analysis
The collected data relating to number of moths, moths’ ratio and eggs produced at different time intervals were used in the statistical analysis. The differences among experimental runs were tested first, which were non-significant. Therefore, data of both runs were pooled. Normality in the data was tested by Shapiro-Wilk normality test (Shapiro and Wilk, 1965), which indicated that data were normally distributed. Therefore, original data were used in the statistical analysis. Two-way analysis of variance (ANOVA) was used to infer the significance in the data (Steel et al., 1997). Least significant difference post hoc test at 5% probability was used to observe the differences among means where ANOVA indicated significant differences. Same statistical procedure was repeated for the economic analysis data obtained after computing the costs incurred on harvesting 100 g eggs. Pearson correlation coefficient was computed among number of moths, moths’ ratio and number of eggs produced by different diet thickness and egg densities’ combinations. All statistical computations were done on SPSS statistical software version 19 (IBM, 2012).
2.6 Economic analysis
The costs incurred to harvest 100 g eggs were computed based on the quantity of eggs harvested from each combination. Total number of experiments required for harvesting 100 g eggs were computed based on harvested eggs from each treatment combination. Afterwards, diet and culture cost, number of containers required to execute the experiments, container cost, and labor cost were summed up to compute the total costs incurred on the harvesting of 100 g eggs. Afterwards, the most economic combination was determined based on statistical analysis.
3 Results
3.1 Impact of different diet thicknesses and egg densities on moth production
Different diet thicknesses, egg densities and their combination significantly altered the number of moths emerged at different weeks as well as total number of moths emerged after 4 weeks (Table 1). However, diet thickness has non-significant impact on moths’ ratio at 1st and 2nd week, whereas egg density had non-significant impact on moths’ ratio at 3rd and last two weeks. The highest number of moths were emerged on thin and medium diet, whereas thick diet resulted in the lowest number of total moths were emerged from thick diet (Table 2). Similarly, the highest and the lowest number of total moths were emerged from high and low egg density, respectively (Table 2). Regarding interactions, thick diet and high egg density recorded the highest number of moths, whereas combination of low egg density with thin and thick diets resulted in the lowest number of total moths (Table 2). DF = degree of freedom, SS = sum of squares, MS = mean squares. Means sharing the same letters within a column are statistically non-significant (p > 0.05), NS = non-significant, LSD = least significant difference.
Treatment
Total number of moths
Moths’ ratio (1st week)
DF
SS
MS
P value
DF
SS
MS
P value
Diet thickness (D)
2
114425.20
57212.60
0.0001*
2
12.57
6.28
0.4020NS
Egg density (E)
2
11993601.73
5996800.87
0.0001*
2
48.14
24.07
0.0382*
D × E
4
195361.47
48840.37
0.0001*
4
130.85
32.71
0.0031*
Number of moths (1st week)
Moths’ ratio (2nd week)
Diet thickness (D)
2
85188.04
42594.02
0.0001*
2
3.30
1.65
0.7821NS
Egg density (E)
2
7176964.44
3588482.22
0.0001*
2
60.68
30.34
0.0173*
D × E
4
77660.36
19415.09
0.0001*
4
151.60
37.90
0.0012*
Number of moths (2nd week)
Moths’ ratio (3rd week)
Diet thickness (D)
2
1106.71
553.36
0.3142NS
2
10.04
5.02
0.0015*
Egg density (E)
2
505362.31
252681.16
0.0001*
2
3.69
1.84
0.0701NS
D × E
4
33326.49
8331.62
0.0001*
4
4.75
1.19
0.01415*
Number of moths (3rd week)
Moths’ ratio (4th week)
Diet thickness (D)
2
747.91
373.96
0.0003*
2
0.84
0.42
0.0560*
Egg density (E)
2
1550.04
775.02
0.0001*
2
0.88
0.44
0.0485*
D × E
4
438.09
109.52
0.0316*
4
2.09
0.52
0.0098*
Number of moths (4th week)
Moths’ ratio (last 2 weeks)
Diet thickness (D)
2
275.20
137.60
0.0249*
2
5.89
2.94
0.0162*
Egg density (E)
2
1149.73
574.87
0.0001*
2
2.42
1.21
0.1636NS
D × R
4
591.87
147.97
0.0053*
4
6.75
1.69
0.0487*
Number of moths
Total
1st week
2nd week
3rd week
4th week
Diet thickness
Thin (T)
851.67 a
672.27 a
157.67
17.40 a
4.80b
Medium (M)
752.27 a
587.93b
149.73
10.60b
4.00b
Thick (Th)
865.47b
686.53 a
161.67
7.67b
9.60 a
LSD 0.05
33.46
26.77
NS
4.48
4.29
Egg density
Low (L)
271.87c
219.13c
46.27c
5.27c
1.67b
Medium (Md)
684.20b
546.47b
123.33b
10.87b
3.53b
High (H)
1513.33 a
1181.13 a
299.47 a
19.53 a
13.20 a
LSD 0.05
33.46
26.77
15.93
4.48
4.29
Diet thickness × Egg density
T × L
311.00f
252.80f
49.80f
7.40 bcd
2.40 bc
T × Md
818.20c
630.00c
169.80c
13.60b
4.80 bc
T × H
1425.80b
1134.00b
253.40b
31.20 a
7.20 bc
M × L
205.80 g
160.40 g
39.20f
5.20 cd
1.00c
M × Md
576.20 e
475.80 e
86.20 e
11.80 bc
2.40 bc
M × H
1474.80b
1127.60b
323.80 a
14.80b
8.60b
Th × L
298.80f
244.20f
49.80f
3.20 d
1.60 bc
Th × Md
658.20 d
533.60 d
114.00 d
7.20 bcd
3.40 bc
Th × H
1639.40 a
1281.80 a
321.20 a
12.60 bc
23.80 a
LSD 0.05
57.96
46.37
27.59
7.76
7.43
Thick diet, high egg density and combination of thick diet and high density resulted in the highest number of moths at 1 week after the initiation of the experiment. The lowest number of moths at the end of 1st week were recorded for medium diet, low egg density and combination of thin and thick diet with low egg density. Diet thickness had non-significant impact on number of moths at the end of 2nd week, whereas the highest and the lowest number of moths were recorded for high and low egg density respectively at the end of 2nd week (Table 2).
Interestingly, thin diet recorded the highest number of moths at the end of 3rd week, while remaining diet thicknesses resulted in similar number of moths. The highest and the lowest number of moths were recorded for high and low egg density, respectively. Regarding interaction, thin diet with high density resulted in the highest number of moths at the end of 3rd week, while thick diet and low egg density combination recorded the lowest number of moths at the end of 3rd week. The highest number of moths were noted for thick diet and high egg density at the end of 4th week, whereas remining diet thicknesses and egg densities produced similar number of moths. Regarding interactions, thick diet with high egg density recorded the highest number of moths at the end of 4th week, while medium diet and low egg density combination resulted in the lowest number of moths (Table 2).
The moths’ ratio was not affected by different diet thicknesses during 1st two weeks of the study (Table 3). Means sharing the same letters within a column are statistically non-significant (p > 0.05), NS = non-significant, LSD = least significant difference.
Moths’ ratio (%)
1st week
2nd week
1st two weeks
3rd week
4th week
last 2 weeks
Diet thickness
Thin (T)
79.30
18.04
97.34b
2.05 a
0.63 ab
2.68 a
Medium (M)
79.08
18.53
97.61 ab
1.90 a
0.50b
2.39 ab
Thick (Th)
80.29
17.90
98.19 a
0.98b
0.83 a
1.81b
LSD 0.05
NS
NS
0.59
0.59
0.27
0.59
Egg density
Low (L)
80.40 a
17.00b
97.40
2.01 a
0.61 ab
2.62
Medium (Md)
80.17 a
17.73b
97.89
1.60 ab
0.51b
2.16
High (H)
78.10b
19.74 a
97.84
1.32b
0.84 a
2.11
LSD 0.05
1.92
1.91
NS
0.59
0.27
NS
Diet thickness × Egg density
T × L
81.31 ab
15.61 de
96.92b
2.36 a
0.79b
3.14 a
T × Md
76.95c
20.83 ab
97.78 ab
1.63 abc
0.58b
2.22 ab
T × H
79.64 abc
17.69 bcde
97.32b
2.17 a
0.51b
2.68 a
M × L
78.24 bc
18.66 abcd
96.89b
2.62 a
0.49b
3.11 a
M × Md
82.51 a
15.00 e
97.52 ab
2.06 ab
0.42b
2.48 ab
M × H
76.48c
21.93 a
98.41 a
1.00c
0.58b
1.59b
Th × L
81.66 a
16.73 cde
98.39 a
1.07 bc
0.54b
1.61b
Th × Md
81.04 ab
17.35 cde
98.39 a
1.10 bc
0.51b
1.61b
Th × H
78.18 bc
19.62 abc
97.79 ab
0.78c
1.43 a
2.21 ab
LSD 0.05
3.32
3.31
1.02
1.02
0.46
1.02
However, low, and medium egg density during 1st week and high egg density during 2nd week resulted in the highest moths’ ratio. Regarding interaction, medium diet and medium egg density and thick diet with low egg density produced the highest number of moths at the end of 1st week, whereas medium diet with high egg density resulted in the lowest moth ratio. Similarly, the highest moth ratio was recorded for medium diet and high egg density combination, whereas medium diet and medium egg density combination resulted in the lowest moths’ ratio at the end of 2nd week (Table 3). The first two weeks resulted in 97–98% of the total moths emerged during the experiment indicating that harvesting/collecting moths only two weeks is sufficient, and the cost incurred on remaining two weeks can be reduced by terminating the experiments at the end of 2nd week.
3.2 Impact of different diet thicknesses and rearing densities on egg production
The individual and interactive effects of diet thicknesses and egg densities significantly affected the egg production at 3rd and 5th days as well as total number of eggs (Table 4). DF = degree of freedom, SS = sum of squares, MS = mean squares.
Treatment
Egg collection (3rd day)
DF
SS
MS
P value
Diet thickness (D)
2
1194275.37
597137.68
0.0001*
Egg density (E)
2
49279291.45
24639645.73
0.0001*
D × E
4
1686356.65
421589.16
0.0001*
Egg collection (5th day)
Diet thickness (D)
2
113406.00
56703.00
0.0001*
Egg density (E)
2
405104.11
202552.06
0.0001*
D × E
4
215990.85
53997.71
0.0001*
Total egg collection
Diet thickness (D)
2
356116.66
178058.33
0.0045*
Egg density (E)
2
79814210.09
39907105.04
0.0001*
D × E
4
952342.59
238085.65
0.0001*
Thin and medium diet resulted in the highest number of eggs produced at the end of 3rd day, while thick diet resulted in the lowest number of eggs (Table 5). Similarly, the highest and the lowest number of eggs were collected from high and low egg density, respectively at the end of 3rd day. Regarding interactions, thin diet with high egg density combination resulted in the highest number of eggs, while thick diet with low egg density resulted in the lowest number of eggs at the end of 3rd day. Means sharing the same letters within a column are statistically non-significant (p > 0.05), NS = non-significant, LSD = least significant difference.
Egg collection (mg)
3rd day
5th day
Total
Diet thickness
Thin (T)
1658.05b
342.47 a
2000.52 a
Medium (M)
1988.69 a
369.98 a
2358.67 a
Thick (Th)
2016.85 a
252.43b
2269.28b
LSD 0.05
79.48
29.28
124.53
Egg density
Low (L)
627.51c
192.66c
820.17c
Medium (Md)
1846.27b
354.03b
2200.30b
High (H)
3189.81 a
418.19 a
3608.00 a
LSD 0.05
79.48
29.28
124.53
Diet thickness × Rearing density
T × L
574.39 g
229.50 ef
808.89 ef
T × Md
1959.58 d
280.67 cd
2240.25 d
T × H
3516.58 a
247.13 de
3763.71 a
M × L
544.67 g
193.88 fg
738.55f
M × Md
2069.56 d
453.10b
2522.66c
M × H
3351.85b
462.96b
3814.81 a
Th × L
763.47f
154.60 g
918.07 e
Th × Md
1509.66 e
328.32c
1837.98 d
Th × H
2701.01c
544.48 a
3245.49b
LSD 0.05
137.67
50.71
215.70
Medium and thick diets produced the highest number of eggs at the end of 5th day, whereas thin diet produced the lowest number of eggs. High and low egg densities resulted in the highest and the lowest number of eggs at the end of 5th day, respectively (Table 5). Regarding interaction, thick diet with high egg density recorded the highest eggs production at the end of 5th day, whereas thick diet with low egg density resulted in the lowest egg production (Table 5).
Medium and thick diets and high egg density resulted in the highest total egg production, while low density and thin diet noted the lowest total egg production. The combination of thin and medium diet with high egg density resulted in the highest egg production, while combination of thick diet with low egg density resulted in the lowest total egg production (Table 5).
3.3 Correlation among number of moths, moths’ ratio, and egg production
Different life history traits of MFM, i.e., number of moths, moths’ ratio and egg production exhibited significant positive and negative correlations among each other. Total egg production has strong positive correlation with number of moths emerged at the end of 1st week (Fig. 1). The number of moths emerged at the end of 2nd week had strong positive correlations with total number of moths, total number of eggs, number of eggs produced at the end of 3rd and 5th days, and number of moths emerged at the end of 1st week. The moths’ ratio at the end of first week has strong negative correlation with the moths’ ratio of 2nd week (Fig. 1).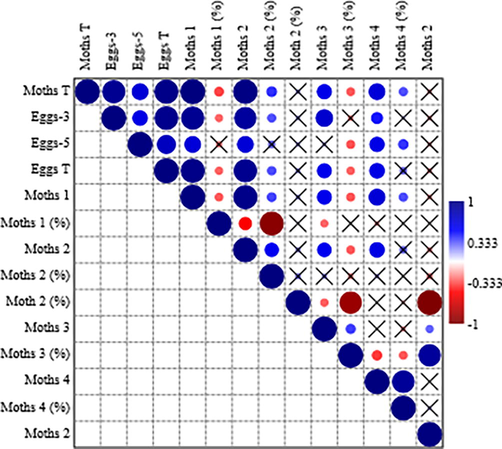
Pearson’s correlation matrix among different life stages of Ephestia Kuehniella reared with different diet thicknesses and rearing densities. Here; Moths T = total number of moths, Eggs 3 = number of eggs collected at 3rd day, Eggs 5 = number of eggs collected at 5th day, Eggs T = total number of eggs, Moths 1 = number of moths observed at the end of 1st week, Moths 1 (%) = ratio of the moths during 1st week, Moths 2 = number of moths observed at the end of 2nd week, Moths 2 (%) = ratio of the moths during 2nd week, Moths 3 = number of moths observed at the end of 3rd week, Moths 3 (%) = ratio of the moths during 3rd week, Moths 4 = number of moths observed at the end of 4th week, Moths 4 (%) = ratio of the moths during 4th week, Moths first 2 (%) = ratio of the moths during first two weeks Moths last 2 (%) = ratio of the moths during last two weeks.
3.4 Economic costs of 100 g egg production
Individual and interactive effects of diet thicknesses and egg densities significantly altered number of experiments and containers required to produce 100 g MFM eggs, and cost incurred on diet, eggs, labor, containers, and total cost (Table 6). DF = degree of freedom, SS = sum of squares, MS = mean squares.
Treatment
Total number of experiments required
Total diet and culture cost
DF
SS
MS
P value
DF
SS
MS
P value
Diet thickness (D)
2
12.58
6.29
0.0037*
2
2877.30
1438.65
0.0001*
Rearing density (E)
2
1004.39
502.19
0.0001*
2
1248.04
624.02
0.0001*
D × E
4
32.63
8.16
0.0001*
4
268.16
67.04
0.0001*
Total number of containers required
Total container cost
Diet thickness (D)
2
1258.37
629.19
0.0037*
2
462.21
231.11
0.0037*
Rearing density (E)
2
100438.96
50219.48
0.0001*
2
36892.18
18446.09
0.0001*
D × E
4
3262.68
815.67
0.0001*
4
1198.41
299.60
0.0001*
Labor cost
Total cost
Diet thickness (D)
2
1039.98
519.99
0.0037*
2
1575.95
787.98
0.0364*
Rearing density (E)
2
83007.41
41503.70
0.0001*
2
265462.80
132731.40
0.0001*
D × E
4
2696.43
674.11
0.0001*
4
9115.33
2278.83
0.0002*
Thin and medium diets and low egg density required the highest number of experiments to produce 100 g MFM eggs. Similarly, combination of thin diet and low egg density required the highest number of experiments, whereas combinations of medium and thick diet with high egg density required the lowest number of experiments to produce 100 g MFM eggs (Table 7). Thick diet and low egg density, and combination of thick diet and low egg density incurred the highest diet and culture cost, whereas thin diet, high egg density and combination of thin diet with medium and high egg density resulted in the lowest culture and diet cost. Thin and medium diets and low egg density required higher number of containers and incurred higher container and labor cost compared to the rest of diet thicknesses and egg densities. Regarding interactions, thin and medium diet in combination with low egg density incurred the highest cost on container and labor, while all diet thicknesses with high egg destiny recorded the lowest cost in this regard. Medium diet and low egg density incurred the highest total cost on 100 g egg production, while thin diet and high egg density resulted in the lowest total cost. Regarding interactive effect, medium diet with low egg density incurred the highest total cost on 100 g egg production, while thin and medium diet with high egg density recorded the lowest cost on the production of 100 g MFM eggs. Although egg production revealed that the combination of thick diet and high egg density was more feasible, economic analysis indicated that high egg density should be combined with thin or medium diet for economic mass production of MFM eggs. Here, Exp 100 = total number of experiments required to collect 100 mg eggs, TDCC = total diet and culture cost, Means sharing the same letters within a column are statistically non-significant (p > 0.05), LSD = least significant difference.
Exp 100
TDCC (US$)
Total number of containers required
Container cost (US$)
Labor cost (US$)
Total cost (US$)
Diet thickness
Thin (T)
7.06 a
11.74c
70.62 a
42.80 a
64.20 a
118.74b
Medium (M)
7.22 a
23.35b
72.21 a
43.76 a
65.64 a
132.76 a
Thick (Th)
6.03b
31.21 a
60.28b
36.53b
54.80b
122.54 ab
LSD 0.05
0.72
1.94
7.25
4.39
6.59
12.83
Rearing density
Low (L)
13.35 a
29.21 a
133.54 a
80.94 a
121.40 a
231.55 a
Medium (Md)
4.46b
20.46b
44.61b
27.04b
40.56b
88.05b
High (H)
2.50c
16.63c
24.95c
15.12c
22.68c
54.44c
LSD 0.05
0.73
1.95
7.26
4.40
6.60
12.84
Diet thickness × Rearing density
T × L
14.02 a
15.93 d
140.16 a
84.95 a
127.42 a
228.29b
T × Md
4.53c
10.29 e
45.27c
27.44c
41.16c
78.88 e
T × H
2.64 d
9.01 e
26.43 d
16.02 d
24.03 d
49.06f
M × L
15.04 a
34.17 ab
150.35 a
91.12 a
136.68 a
261.98 a
M × Md
4.09c
18.60 d
40.92c
24.80c
37.20c
80.59 e
M × H
2.54 d
17.29 d
25.35 d
15.37 d
23.05 d
55.70f
Th × L
11.01b
37.54 a
110.12b
66.74b
100.11b
204.39c
Th × Md
4.77c
32.49b
47.65c
28.88c
43.32c
104.69 d
Th × H
2.31 d
23.59c
23.07 d
13.98 d
20.97 d
58.55 ef
LSD 0.05
1.25
3.38
12.55
7.61
11.41
22.23
4 Discussion
Egg production of MFM was significantly altered by different diet thicknesses and egg densities used in the study. The thin and medium diet thickness and high egg density recorded the highest eggs production compared to the rest of the treatments. Similarly, combination of thin and medium diet thickness with high egg density resulted in the highest egg production, whereas combination of medium and thick diet with low egg density gave poor egg production. The economic analysis revealed that thin diet and high egg density were economic to produce 100 mg eggs. Similarly, combination of thin and medium diet with high egg density proved the most economic combination compared to the rest of the treatments in the study. The results validated our hypothesis that diet thickness and egg density pose significant impacts on egg production of MFM. Similarly, the study has identified the most economic combinations of diet thickness and egg density to produce MFM eggs. Nevertheless, moths’ ratio and egg production also indicated that the experiments for producing moths should be terminated at the end of 2nd week (egg harvesting should be terminated two weeks after the 1st moth was emerged) and egg collection should be terminated at 4 days to cut down the labor and other related expenses. >90% of the moths were harvested within two weeks after the emergence of first moth (Table 3). Similarly, 80–90% of eggs were harvested 3 days after the moths were placed in egg laying (Table 5). Therefore, harvesting eggs for 1 time only at 4th day would significantly reduce the labor and operating costs and 90–100% eggs will be harvested. Numerous earlier studies have reported that different diets had significant impact on the life history traits of MFM (Faal-Mohammad-Ali and Shishehbor, 2013; Kurtuluş et al., 2020; Tarlack et al., 2015). However, these studies did not investigate the economics of the mass production of MFM. The mass rearing should be economic to compete with the other prevalent pest management options. The economic mass rearing of biological control agents is dependent on the economic mass rearing of their hosts. Biological control method can only compete with other pest management methods if the biological control agents are reared with minimum possible cost. The most economic treatment of the study incurred 49 USD on the rearing of 100 g MFM eggs, while these are available for 330 USD in the market (www. greenmethods.com). This indicates that the study could help to reduce the egg cost by 7 times of the market price.
Higher egg production in high density treatment is directly linked to higher number of eggs used for mass rearing. The higher number of eggs resulted in more moth production, which laid more eggs compared to low density treatments. The thick diet, although resulted in a greater number of moths, it can be considered economic due to higher costs involved. It is thought that thin and medium diet provide sufficient nutritional requirements of the emerging moths; thus, providing more diet yields nothing special. However, higher density than 75 mg should be tested in the future studies to infer whether these are more economical than the currently used egg densities for mass rearing. Mamay and Mutlu (2019) optimized container size and rearing larval density for economic and rapid mass production of Oenopia conglobata (Linnaeus, 1758). The authors also reported that using low rearing density is unsuitable due to higher economic costs involved and recommended the use of high rearing density like we found in this study.
The MFM adults emerged after 30–36 days when reared on corn flour, wheat flour and hazelnut flour. Maize flour was regarded more efficient for mass production of MFM (Polat, 2008). Although similar results were obtained for adult emergence in the current study, slight difference can be opted to the medium and diet used in the current study. In another study, MFM adults emerged 35–40 days after the initiation of experiment (Altuntaş et al., 2010).
Alpkent (2009) observed adult emergence at 45–50 days with 2 cm diet thickness. Similarly, Karakuş (2018) used 27×37×7 cm containers and sprinkled 0.1 g eggs and reported adult emergence 35–40 days after the initiation of the experiment. The authors recommended to harvest eggs 3 days after the moths were placed in the egg laying containers. However, their recommendation was not based on the economic costs involved in mass rearing. We also recommend harvesting eggs 3 days after the moths are shifted to egg laying containers due to low economic costs involved in the egg production.
Similarly, Kılınçer (2010) stated that MFM eggs were produced by homogeneously distributing 5000 eggs into 27×37×7 cm tubs containing 750 g wheat flour and bran mixture. The amount of mixture used, size of plastic tub used, and the density of eggs is in line with the results of current study. However, again these recommendations were based on egg quantity and economic costs involved were not computed. Demirtaş (2014) placed 400–500 eggs of MFM on wheat bran, cracked corn and cracked wheat, and the eggs laid by the adults were collected once in 48 h. In this study, two harvests were made on the 3rd and 5th days. The recommendations were to harvest the eggs for both time periods. Our study clarifies that harvesting eggs at 3 days after the moths are placed in egg laying containers is sufficient as > 80% eggs are laid. The second harvest would cost similar amount and would give the rest of 20% eggs. Thus, high egg density, thin and medium diet, terminating moth collection at 2nd week and terminating egg collection at 3 days are the most economic options for the mass production of MFM eggs.
5 Conclusions
The results of the current study indicated that combination of medium and thin diet thicknesses with high egg density are the most economic for mass production of MFM. Interestingly, the study revealed that terminating moths’ collection at 2nd week and egg collection at 3 days after the moths are placed in egg laying containers is the most suitable times for the economic mass rearing of MFM. However, higher gg densities than 75 mg need further testing on thin and medium diet thickness to explore whether higher densities are more economic than the densities recommended in this study.
Acknowledgements
The current study was supported by Scientific Research Projects Commission of Harran University, Şanlıurfa Turkey (Project No:17175). A part of this manuscript was Masters’ thesis of Havva Karakuş. This work was supported by the King Khalid University through a grant KKU/RCAMS/22 under the Research Center for Advance Materials (RCAMS) at King Khalid University, Saudi Arabia.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Reproductive performance and mating behavior of the predatory bug, Anthocoris minki Dohrn, 1860 (Hemiptera: Anthocoridae) Turkish J. Entomol.. 2018;42:117-125.
- [Google Scholar]
- Bazı Bitkisel Uçucu Yağların Ephestia kuehniella’ya (Lepidoptera: Pyralidae) Fümigant Etkileri Üzerinde Araştırmalar. Doktora Tezi. 2009
- [Google Scholar]
- Pesticides driven pollution in Kuwait: The first evidence of environmental exposure to pesticides in soils and human health risk assessment. Chemosphere. 2021;273:129688
- [CrossRef] [Google Scholar]
- The effects of parasitism by the ectoparasitoid Bracon hebetor Say (Hymenoptera: Braconidae) on host hemolymph proteins in the Mediterranean flour moth Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) Turkish J. Zool.. 2010;34:409-416.
- [Google Scholar]
- Learnings from over a decade of increasing pesticide resistance in the redlegged earth mite, Halotydeus destructor (Tucker) Pest Manag. Sci.. 2021;77:3013-3024.
- [CrossRef] [Google Scholar]
- Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol.. 2002;47(1):817-844.
- [Google Scholar]
- Biological control and sustainable food production. Philos. Trans. R. Soc. B Biol. Sci.. 2008;363:761-776.
- [CrossRef] [Google Scholar]
- The status of biological control and recommendations for improving uptake for the future. BioControl. 2018;63:155-167.
- [CrossRef] [Google Scholar]
- E. Borzoui B. Naseri M. Mohammadzadeh-Bidarani Adaptation of Habrobracon hebetor (Hymenoptera: Braconidae) to Rearing on Ephestia kuehniella (Lepidoptera: Pyralidae) and Helicoverpa armigera (Lepidoptera: Noctuidae) J. Insect Sci. 16 1 2016 2016 10.1093/jisesa/iew001 12.
- Yumurta paraziti Trichogramma spp. Hymenopt. Trichogrammatidae)’nin un güvesi 1987:12-14.
- [Google Scholar]
- The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B Biol. Sci.. 2011;366:1987-1998.
- [CrossRef] [Google Scholar]
- The egg parasitoids Trichogramma: from laboratory mass rearing to biological control of lepidopteran pests. Biocontrol Sci. Technol.. 2021;31:661-693.
- [CrossRef] [Google Scholar]
- Demirtaş, Ş.N., 2014. Besinin Trichogramma brassicae Bezdenko ve Trichogramma evanescens Westwood (Hym.; Trichogrammatıdae) türlerinin bazı biyolojik özelliklerine etkisi.
- Crop Losses due to insect pests: Global and Indian Scenario. Indian J. Entomol.. 2015;77:165.
- [CrossRef] [Google Scholar]
- Biological parameters of Bracon hebetor (Hym.: Braconidae) parasitizing Ephestia kuehniella (Lep.: Pyralidae): effect of host diet. J. Crop Prot.. 2013;2:411-419.
- [Google Scholar]
- Atmospheric Occurrence of Organochlorine Pesticides and Inhalation Cancer Risk in Urban Areas at Southeast Brazil. Environ. Pollut.. 2021;271:116359
- [CrossRef] [Google Scholar]
- Effect of Temperature on Parasitism and Host-Feeding of <I>Trichogramma turkestanica</I> (Hymenoptera: Trichogrammatidae) on <I>Ephestia kuehniella</I> (Lepidoptera: Pyralidae) J. Econ. Entomol.. 2002;95:50-56.
- [CrossRef] [Google Scholar]
- Dusak, H., 2018. Biyolojik mücadele makrobiyal ajanı predatör Oenopia conglobata (Linnaeus)(Coleoptera: Coccinellidae)’nın laboratuvar koşullarında kannibalistik davranışlarının belirlenmesi/Determination of cannibalistic behaviors of biological macrobial agent predatory .
- IBM, C., IBM SPSS Inc. 2012. SPSS Statistics for Windows. IBM Corp. Released 2012. Version 20. 1–8.
- Risk of resistance and cross-resistance development to selection with imidacloprid and level of heritability in Oxycarenus hyalinipennis Costa (Hemiptera: Lygaeidae): a potential pest of cotton. Phytoparasitica. 2021;49:287-297.
- [CrossRef] [Google Scholar]
- H. Karakuş Determination of suitable nutrient thickness and egg spreading amount in mass rearing of the mediterranean flour moth, Ephestia kuehniella zeller (Lepidoptera: Pyralidae) 2018 Harran University.
- Kılınçer, N., 2010. Ephestia kuehniella’da Transferin Geninin Moleküler Karakterizasyonu ve Savunma Reaksiyonlarındaki Rolü Üzerinde Araştırmalar.
- Effects of some insecticides used in maize (Zea mays L.) on preimaginal stages of Trichogramma evanescens Westwood (Hymenoptera: Trichogrammatidae). Turkish. J. Entomol.. 2015;39
- [CrossRef] [Google Scholar]
- Influence of different diets on some biological parameters of the Mediterranean flour moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) J. Stored Prod. Res.. 2020;85:101554
- [CrossRef] [Google Scholar]
- Optimizing container size and rearing density for rapid and economic mass rearing of Oenopia conglobata (Linnaeus, 1758) (Coleoptera: Coccinellidae) Turkish J. Entomol. 2019:395-408.
- [Google Scholar]
- Pesticide residues, health risks, and vegetable farmers’ risk perceptions in Punjab, Pakistan. Hum. Ecol. Risk Assess. An Int. J.. 2021;27:846-864.
- [CrossRef] [Google Scholar]
- Current challenges to the implementation of classical biological control. BioControl. 2018;63:1-9.
- [CrossRef] [Google Scholar]
- Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens. Mass Production of Beneficial Organisms: Invertebrates and Entomopathogens; 2014.
- Detection of different densities of Ephestia kuehniella pest on white flour at different larvae instar by an electronic nose system. J. Stored Prod. Res.. 2019;84:101522
- [CrossRef] [Google Scholar]
- Effect of different cold storage periods on parasitization performance of Trichogramma cacoeciae (Hymenoptera, Trichogrammatidae) on eggs of Ephestia kuehniella (Lepidoptera, Pyralidae) Biocontrol Sci. Technol.. 2004;14:441-447.
- [CrossRef] [Google Scholar]
- Occurrence of pesticide residues in fruits and vegetables for the Eastern Mediterranean Region and potential impact on public health. Food Control. 2021;119:107457
- [CrossRef] [Google Scholar]
- Polat, P., 2008. Adult longevity, fecundity and sex ratio on ephestia kuehniella (Zeller, 1879) (lepidoptera: pyralidae) feeds three different larval diet during larval period. Ondokuz Mayis University.
- An extensive review on the consequences of chemical pesticides on human health and environment. J. Clean. Prod.. 2021;283:124657
- [CrossRef] [Google Scholar]
- Formulation of biological control agents. In: Jones D.G., ed. Exploitation of Microorganisms. Dordrecht: Springer Netherlands; 1993. p. :411-439.
- [Google Scholar]
- The Effect of Essential Oils from Laurus nobilis and Myrtus commonis on the Adults of Mediterranean Flour Moth, Ephestia kuehniella Zeller (Lep.: Pyralidae) J. Essent. Oil Bear. Plants. 2014;17:553-561.
- [CrossRef] [Google Scholar]
- Comparison of different European strains of Trichogramma aurosum (Hymenoptera: Trichogrammatidae) using fertility life tables. Biocontrol Sci. Technol.. 2008;18(1):75-86.
- [Google Scholar]
- P. Senthil Kumar G. Janet Joshiba Pesticides Pollution and Analysis in Water 2021 10.1007/978-3-030-54719-6_9 337 349.
- Intraguild predation of Orius niger (Hemiptera: Anthocoridae) on Trichogramma evanescens (Hymenoptera: Trichogrammatidae) Eur. J. Entomol. 2017;114:609-613.
- [Google Scholar]
- An analysis of variance test for normality (complete samples) Biometrika. 1965;52(3-4):591-611.
- [Google Scholar]
- Sönmez, C., Mamay, M., 2018. Biological control in sustainable agriculture, in: Proceedings of the International GAP Agriculture and Livestock Congress. p. 362.
- Steel, R., Torrei, J., Dickey, D. 1997. Principles and Procedures of Statistics A Biometrical Approach. A Biometrical Approach.
- Larvicidal activity of substituted chalcones against <scp> Aedes aegypti </scp> (Diptera: Culicidae) and non-target organisms. Pest Manag. Sci.. 2021;77:325-334.
- [CrossRef] [Google Scholar]
- Life history and fecundity rate of Ephestia kuehniella (Lepidoptera: Pyralidae) on different wheat flour varieties. Arch. Phytopathol. Plant Prot.. 2015;48:95-103.
- [CrossRef] [Google Scholar]
- The impact of resource limitation and the phenology of parasitoid attack on the duration of insect herbivore outbreaks. Theor. Popul. Biol.. 2002;62(3):259-269.
- [Google Scholar]
- Pest Control: Cultural and Environmental Aspects. Bull. Entomol. Soc. Am.. 1980;26:478-479.
- [CrossRef] [Google Scholar]
- Biological control agents for plant pathogens. In: Van Driesche R.G., Bellows T.S., eds. Biological Control. Boston, MA: Springer US; 1996. p. :93-101.
- [Google Scholar]
- Biological control using invertebrates and microorganisms: plenty of new opportunities. BioControl. 2018;63:39-59.
- [CrossRef] [Google Scholar]
- Urbanization and the Viability of Local Agricultural Economies. Land Econ.. 2011;87:109-125.
- [CrossRef] [Google Scholar]
- Influence of larval density on biological fitness of Ephestia kuehniela Zeler (Lepidoptera Pyralidae) New Zeal. Plant Prot.. 2007;60:199-202.
- [CrossRef] [Google Scholar]




