

Translate this page into:
Neuroprotective effect of endophytic fungal antioxidant polyphenols on cerebral ischemic stroke-induced Albino rats; memory impairments, brain damage, and upregulation of metabolic proteins
⁎Corresponding author at: Department of Medical Laboratory Sciences, College of Applied Medical Sciences, Majmaah University, Majmaah, Saudi Arabia. m.palanisamy@mu.edu.sa (Palanisamy Manikandan)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Background
Stroke is one of the leading causes of mortality and disability throughout the world. Recently, antioxidant therapies were attempted to reduce apoptotic cell death in cerebral ischemia animal model.
Purpose
To study the neuroprotective properties of polyphenol derived from fungal endophyte analyzed on experimental Albino rat.
Methods
Polyphenols producing endophytic fungi was initially isolated from the seeds of Moringa oleifera Lam. The endophytes were cultured in potato dextrose broth and the potent strain Simplicillium sp. ED7 produced maximum phenolic content (86.42 ± 5.3 mg GAE/g) than other fungi. Polyphenols were extracted with solvent and used for the determination of neuroprotective properties.
Results
Isoflurane was used to induce stroke in Albino rat and treated polyphenols showed reduced neurological deficits and improved neuroprotective properties. The ischemic Albino rats treated with polyphenols restored memory loss. The increased dosage of polyphenol improved the biosynthesis of more antioxidant enzymes than lower dosages. Central artery occlusion evoked about 2.28-fold increase in reactive oxygen species in brain tissue and the generation of reactive oxygen species was decreased in polyphenol treated animal.
Conclusion
Albino rats treated with different doses of polyphenol had decrease ROS amount than sham group. The elevated level of cytochrome revealed mitochondrial damage in stroke induced control Albino rat. After 24 h of reperfusion on Albino rat, upregulation of total p65 and phospho-p65 were determined. The present finding revealed that polyphenl has a neuroprotective property in ischemia and regulate metabolic enzymes and restore brain injury.
Keywords
Stroke
Cerebral ischemia
Polyphenols
Neuroprotective
Metabolic enzymes
Upregulation

1 Introduction
Ischemic heart disease is one of the leading causes of death throughout the world; therefore many strategies have been developed to protect myocardium and haemostasis. In most of the cases, myocardial ischemia forms when normal blood flow to the myocardium is continuously reduced, either in terms of relative to increased tissue demand or absolute flow rate. The most important problem of diminished blood supply to the myocardium is myocardial injury and is irreversible, which mainly happens when ischemia continues more than 20 min in severe cases (Ferdinandy et al., 2007). Reperfusion is the rapid restoration process at the site of ischemic region of the myocardium, supplies metabolic products and oxygen and remove ischemic metabolites. Myocardial ischemia/reperfusion injury is generally caused by myocardial ischemia and subsequent reperfusion process. Myocardial ischemia/reperfusion is characterized by development of tissue necrosis, increased incidence of arrhythmias decreased myocardial function. The reperfusion process is useful to the benefit of the heart; however it is linked with myocardial injury. Reperfusion of ischemic myocardium results in the development of cellular damage; moreover, the pathogenesis is not clearly understood (Ibáñez et al., 2015).
Reactive oxygen species (ROS) are produced from the cells and the excess amount of ROS is highly toxic to the cells and involved in cellular damages. ROS includes hydroxyl ion, superoxide, hydrogen peroxide, sulfhydryl, nitric oxide, alkoxyl, etc.are generated from the cellular components. ROS is toxic to lipids, proteins, enzymes and nucleic acids and causes necrosis and apoptosis (Al-Dhabi et al., 2020). Oxidative stress plays an important role in the development of various human diseases, including, cardiovascular, Alzheimer's disease, neurodegenerative disorders, cancer, kidney disease and diabetes etc. In cellular system, oxidative stress results the generation of unneutralized active components within the cells. In recent years, various antioxidant molecules have been synthesized and these chemical antioxidants are not safe to the cellular system. Hence, there is a need to screen novel antioxidant compounds against ROS to balance cellular functions (Al-Ansari et al., 2021).
Antioxidants are naturally available chemical compounds with novel therapeutic properties against these chemical and naturally induced oxidative damages. These antioxidants inhibit the generation of free radicals by scavenging the free radical chain reaction at early stage or by binding to the oxidation promoter and this process involved various molecular mechanisms (Atif et al., 2020). Phenolic compounds, ascorbic acids, and thiols are the major antioxidant compounds from the natural sources. Medicinal plants are widely studied for antioxidant properties. Moreover, fungi are gaining much more interest in recent years for the biosynthesis of various novel metabolites. Fungal endophytes have the potential to produce various secondary metabolites. The endophytes produce various bioactive substances and are useful in modern medicine, industry and agriculture and used as anticancer, anti thrombus, antibiotics and antioxidants (Anand et al., 2021).
Endophytes produce various novel molecules, including xanthones, tetralones, terpenoids, steroids, quinones, phenolic acids, flavonoids, chinones, benzopyranones and alkaloids. Among these chemical compounds, flavonoids and phenolic compounds are the major bioactive compounds (Huang et al., 2018). The endophytic fungus Annulohypoxylon stygium produce phenolic acids (Cheng et al., 2014). Endophytes were isolated from various medicinal plants and these are novel sources for the discovery of naturally available products (Huang et al., 2008). Plant endophytic strains have rich species diversity and associated with the taxonomy of host plants and environmental variations (Wu et al., 2013). The diversity of endophytic fungi influenced on the production of diverse bioactive compounds. In optimized culture conditions, endophytic fungi produce various bioactive metabolites. Therefore, endophytic fungi are not only a novel source for the development of cardiovascular therapeutic molecules, but also a bioreactor for the production of novel lead molecules more economically and easily.
Moringa oleifera Lam. is widely used for medicine and nutrient purposes. It has various health benefits, including anti-cholesterol, antifungal, antibacterial, antioxidant, anti-inflammatory, pain relief, anti-ulcer, would healing and immunomodulatory functions (Amina et al., 2019). The antibacterial, antifungal, and antioxidant properties of medicinal plant have been studied previously using various experimental models. However, only few studies on the endophytic strain and these naturally occurring endophytic fungi resources should be utilized for biological functions. In this study, the bioactive molecules were analyzed in vivo using animal model and the role of stoke preventing activity was analyzed.
2 Materials and methods
2.1 Endophytic fungi
The young seeds of M. oleifera were collected and separated from the plant. The seed was cut into small pieces and soaked in ethanol (70 %, v/v) for two minutes, followed by sodium hypochlorite (5 %) treatment for five minutes. It was further washed with ethanol for 60 s. The pre-treated seed was placed in potato dextrose agar plates which were amended with kanamycin sulphate (5 mg/100 mL) and ampicillin (5 mg/100 mL). These plates were incubated for 8–10 days at 28 ± 1 °C. The morphologically different fungi were further purified using the same potatoes dextrose medium and were stored at 2–8 °C in the Laboratory (Baazeem et al., 2021).
2.2 Polyphenol-producing endophytic fungi
The isolated endophytic fungi were cultured into potato dextrose broth medium for 8 days at 28 ± 1 ͦC in an orbital shaker incubator at 150 rpm/min. After 8 days incubation, the culture was filtered using multilayer cheesecloth and the filtrate was obtained. Colour reaction-method was used for the determination of polyphenol (Yang et al., 2015). Briefly, 0.5 mL of culture supernatant was mixed with 0.1 mL chromogenic agent. Chromogenic agent was prepared by mixing FeCl3and K3[Fe(CN)6] in a very glassware. It was mixed and incubated for 10 min at room temperature. The development of blue colour indicated the presence of polyphenols.
2.3 Cultural, morphological and molecular characterization of endophyte
The morphological characters, pigmentation, hyphae, mycelium colour, colony morphology, spore morphology, spore-arrangement, and 18S rDNA sequencing analysis were performed. To determine the colony morphology the selected strain ED7 was cultured on potato dextrose agar and examined under an inverted microscope. Genomic DNA of the endophytic strain ED7 was isolated by standard method and the ITS region was amplified using the ITS1-forward (TCCGTAGGTGAACCTGCGG) and reverse (TCCTCCGCTTA TTGATA TGC) primers (White et al., 1990). PCR reaction mixture and 18S rDNA sequencing were performed as described previously earlier. The final PCR products were sequenced and the sequences were compared using BLAST search tool and the final sequences were submitted in GenBank database (Ding et al., 2013).
2.4 Crude extract preparation
Endophytic strain ED7 was cultured in potato dextrose broth medium and the cell free extract was used as the sample until otherwise stated. The cell free broth was fractionated with petroleum ether, n-butanol and chloroform, individually. The crude extract was concentrated after lyophilisation. It was separated in Thin Layer Chromatography and the phenolic factions were collected and pooled. The solid material was dissolved in minimum volume of water and bioactive potentials were studied.
2.5 Total phenolic content
Total phenolic content of the sample was determined from the extract using Folin–Ciocalteu method. Briefly, 0.2 mL of solvent extract was mixed with 0.8 mL double distilled water in a clean test tube. To this 0.6 mL of diluted Folin–Ciocalteu reagent (1:1 dilution) was added and incubated for 5 min. To the reaction mixture 0.5 M sodium carbonate (1 mL) was added and the colour development was observed at 730 nm against reagent blank. Gallic acid was prepared at various concentrations and the amount of phenolic content was expressed in mg equivalent in gallic acid. Experiments were performed in triplicates and an average value was considered for data processing (Srivastava et al., 2012).
2.6 Experimental animal
Adult Albino rat, 15 weeks old with weight 100 – 110 g were applied in our study. The experimental animals (n = 24) were maintained at 12/12 h dark/light photo period, with 60 % humidity at 30 ± 1 ͦ C. All experimental and control animal were allowed free access to food and water. All standard protocols were followed and ethical approval was obtained from the institutional ethical committee for animals (Ethical Approval Number: 2021/STR/Alb/108-C).
2.7 Induction of ischemic stroke in experimental animal
In this study, ischemia was induced in an experimental animal. The male Albino rats were anesthetized by standard method using isoflurane. The healthy animals were randomly selected and divided into five different groups. The experimental groups were sham group, ischemic stroke group treated with vehicle, stroke group treated with 100, 150 and 200 mg/kg polyphenol. The phenolic compounds were administered once daily and reconstituted with saline. Stroke was induced after two weeks of experimental procedure. Albino rat was anesthetized using chloral hydrate (500 mg/kg) using intraperitoneal injection. An external carotid artery was exposed and a nylon suture was introduced into the lumen of external carotid artery. After 60 min of treatment, blood flow was restored and the suture was withdrawn. The experimental animal retains stability after two weeks of stroke development. Phenolic compound was administered at a single dose/per day for 21 days. In the sham group, all experimental procedures were same except the administration of phenolic compounds.
2.8 Analysis of neuroprotective effect
The neuroprotective effect of polyphenol after stroke was evaluated for Albino rat after 24 h ischemia. Neuroprotective effect was evaluated using various parameters, including symmetry of movements, spontaneous activity and climbing movement of Albino rat. Each test consists of three points and score was analyzed. The maximum allotted scores were 15 (3 × 5).
2.9 Brain tissue for in vitro bio-assays
Cortical tissues were used for the determination of enzyme activity and lipid peroxidation analysis. Cortical tissues were dissected from the experimental and control animal and immediately placed in ice cold condition. It was homogenized with 0.5 mL saline (0.9 %) and centrifuged at 4000 rpm for 10 min. The insoluble matters were removed and the clear supernatant was stored at −20 °C.
2.10 Superoxide dismutase activity
Superoxide dismutase (SOD) was analyzed from the sample as described earlier (Misra and Fridovich, 1972). The extract was mixed with carbonate buffer containing 0.1 mM EDTA and the reaction was performed with 30 mM epinephrine prepared in 0.05 % acetic acid. The rate of epinephrine autooxidation was analyzed at 480 nm for three minutes using UV–Visible spectrophotometer. One SOD unit is defined as the amount of sample required to inhibit the autoxidation of epinephrine about 50 %.
2.11 Total glutathione peroxidase (GPx) activity
The amount of glutathione peroxidase (GPx) activity was evaluated as described earlier (Somani and Husain, 1997). Briefly, the reaction mixture was prepared by adding 200 μl of 1.5 mM NADPH, 200 μl of 0.01 M GSH, 50 mM phosphate buffer and 200 μl of 0.24 units GR. To this reaction mixture, 0.1 mL tissue extract was added and incubated for 10 min at 37 °C. Then, 200 μl 12 mM t-butyl hydroperoxide was added and the absorbance was read at 340 nm for three min and NADPH oxidation was determined. One GPx activity was expressed as the amount of enzyme required to oxidize 1 mM NADPH/min under standard assay condition.
2.12 Lipid peroxidation analysis
Lipid peroxidation was evaluated by assaying MDA levels. MDA levels were evaluated as described previously by the thiobarbituric acid (TBA) method with minor modifications. Sample (0.2 mL) was incubated with 0.2 mL of 0.37 % TBA which was prepared in 50 mM NaOH and 0.2 mL TCA was added and placed in a boiling water bath for 20 min. The final pink chromogen was detected at 532 nm against reagent black using a UV–Visible spectrophotometer. The final result was expressed as μM MDA/g of tissue (Spickett et al., 2010).
2.13 Preparation of cortical mitochondria
Mitochondria were separated from the dissected brain tissue using a commercial kit according to the manufactures instruction. Briefly, the brain was dissected and homogenized using isolation buffer. The homogenized fraction was centrifuged at 2000 rpm for 4 min at 4 °C and the pellet consists of mitochondrial fraction. It was further resuspended in lysis buffer and the supernatant was again centrifuged for 10 min at 14000 rpm and cytosolic fraction was obtained.
2.14 Reactive oxygen species analysis (ROS)
The ROS analysis was performed in cortical tissues as described previously by Wang et al. (Zhang et al., 2010). The ROS level in experimental group was compared with control group (sham).
2.15 Determination of Caspase-3 activity
The amount of caspase-3 activity was analyzed after ischemia and reperfusion injury. Briefly, the cortical tissues were lyzed and centrifuged at 10000 rpm for 4 °C. The supernatant faction was used for the determination of caspase-3 activity. The sample (0.1 mL) was incubated with 2 mM substrate (Ac-DEVD-pNA) and the final absorbance was read at 405 nm using a UV–Visible spectrophotometer.
2.16 Western blot analysis
The processed mitochondria obtained from the cortical tissues were used for the determination of expression of proteins. The proteins were separated by sodium dodecyl sulphate–polyacrylamide gel electrophoresis (SDS-PAGE). The discontinuous SDS-PAGE consists of 11.5 % separating gel and 5.5 % stacking gel with constant voltage (50 V) for 3 h. After electrophoresis, the SDS-PAGE was transferred onto polyvinylidene fluoride (PVDF) membranes by electro blotting method. The PVDF membrane was blocked with 5 % BSA prepared in Tris buffer containing tween-20 (0.1 %) for 1 h. It was washed and incubated with primary antibody at 4 ͦC. The plate was washed three times with tris buffered saline and incubated with secondary antibody. Then the blots were visualized and the expression level was determined.
2.17 Statistical analysis
Data obtained in this study was expressed as mean ± SD. Analysis of variance (ANOVA) was performed to analyze the significant variation and the p value ≤ 0.05 was considered as significant. Duncan and Pearson’s analysis was performed to determine the significant level among control and experimental groups.
3 Results
3.1 Isolation of endophytic fungi for polyphenol production
A total of 13 endophytic fungal strains were isolated from the seed of M. oleifera. The morphologically different fungal strains were cultured in potato dextrose broth medium. Initial screening revealed that only seven endophytic strains have the ability to produce polyphenols. The strain ED7 showed maximum colouration (dark blue) indicated potential producer of polyphenols. The phenol positive fungal strains were subjected for the determination of total phenolic content.
3.2 Determination of phenolic content
Total phenolic content of fungal broth was determined and the amount of phenolic content varied widely. The total phenolic content ranged from 3.592 ± 2.1 mg GAE/g to 86.42 ± 5.3 mg GAE/g. The strain ED7 showed maximum phenolic content (86.42 ± 5.3 mg GAE/g) extracted with petroleum ether. N-butanol showed 56.35 ± 2.7 mg GAE/g, whereas, chloroform possessed reduced amount of phenol (49.47 ± 3.7 mg GAE/g). The strain ED4 showed 38.2 ± 1.5 mg GAE/g, 69.1 ± 4.8 mg GAE/g, and 54.3 ± 2.9 mg GAE/g in n-butanol, petroleum ether and chloroform, respectively. The phenolic content of endophytic fungal strains was described in Table 1.
Strains
Polyphenol (mg GAE/g)
Petroleum ether
n-butanol
Chloroform
ED1
14.2 ± 1.1
40.2 ± 2.4
39.5 ± 2.8
ED2
0.2 ± 0.1
4.2 ± 1.1
2.9 ± 0.1
ED3
0 ± 0
0 ± 0
0.5 ± 0
ED4
69.1 ± 4.8
38.2 ± 1.5
54.3 ± 2.9
ED5
3.592 ± 2.1
3.02 ± 0.5
2.6 ± 0.4
ED6
49.3 ± 4.9
16.2 ± 0.9
18.1 ± 1.5
ED7
86.42 ± 5.3
56.35 ± 2.7
49.47 ± 3.7
3.3 Characterization of polyphenol (PP) producing endophyte ED7
The potent endophytic fungal strain was grown well in potato dextrose agar and reached about 25–29 mm colony diameter within two weeks of incubation at 28 °C. It was convex and detected aerial mycelium, yellowish to pale brown colour and did not produce any soluble pigment. Conidia are very small, cylindrical, smooth walled and octahedral crystals absent. Based on colony morphology and 18S rDNA gene sequencing, the strain ED7 was identified as Simplicillium sp. ED7.
3.4 Neuroprotective role of polyphenol in Albino rat
Albino rat associated with cerebral artery occlusion showed neurological deficits after one day of ischemia. The neurological defect was compared with control animal (Sham-group). The artery occlusion animals showed decreased ability to respond to various stimuli and observed decreased motor ability. Animal treated with PP showed reduced neurological deficits and improved neuroprotective properties. At higher concentrations of PP increased motor activity was observed and the result was illustrated in Fig. 1. In sham group of animals, the score was 14.5 ± 1.3 and it was reduced in central artery occlusion (CAO) experimental group (9.2 ± 1.2). The score increased at higher doses of PP.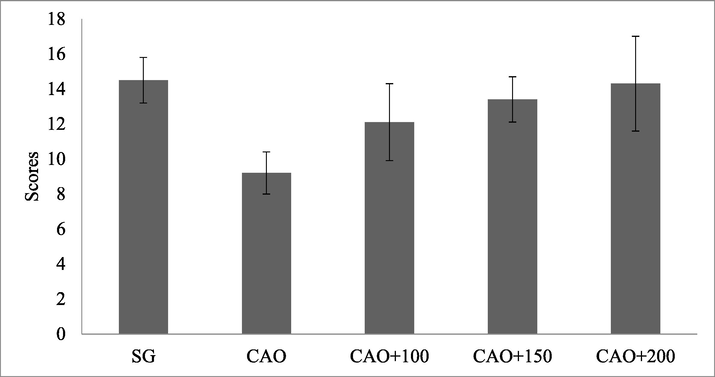
PP treatment decreases ischemia-induced nerve deficits in Albino rat. Albino rat was treated with PP (100 mg. 150 mg, and 200 mg/kg).
3.5 Effect of PP in locomotor activity in experimental animal
The locomotor activity was evaluated in sham group and CAO group, CAO group treated with PP. Locomotor activity was evaluated after 24 h of ischemia and observed significant changes in locomotion. Experimental animals treated with PP at higher concentrations (150/kg and 200/kg) improved locomotor activity. Likewise, rearing activity was also restored by the supplemented PP and increased activity was observed at higher concentrations of PP in the treated experimental group (Fig. 2A and B).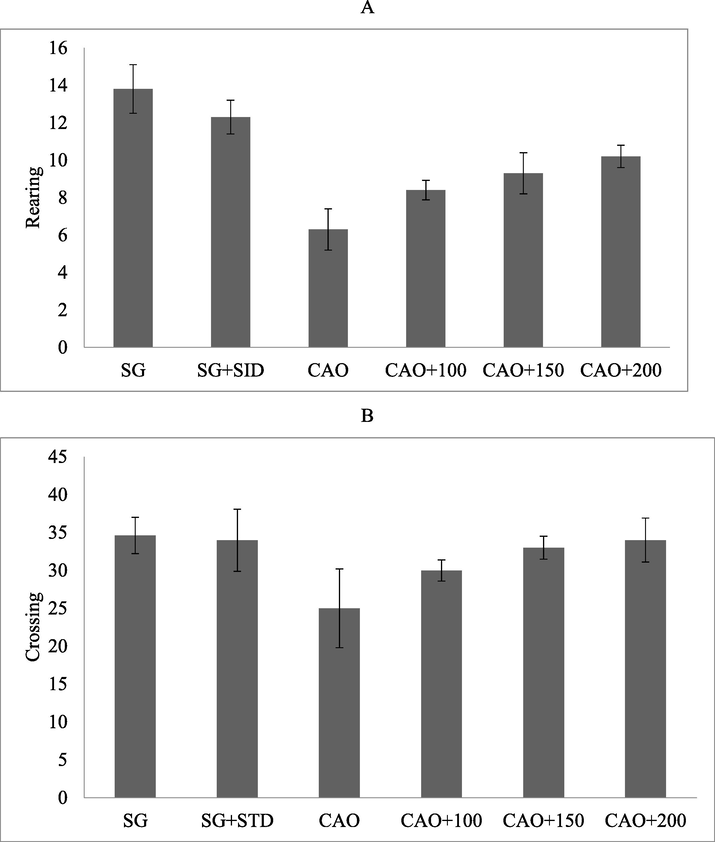
PP treatment of artery occlusion altered motor activity. Albino rat was administered with PP at 100–200 mg/kg via intraperitoneal injection. The result was expressed as number of rearing (A) and number of crossing (B).
3.6 Effect of PP on memory
The experimental animal which was subjected to ischemia was affected by memory loss. In SO group 67.2 ± 1.2 % working memory was registered and SS + standard PP retained 65.8 ± 1.2 % working memory. The PP administered artery occlusion animals showed reduced deficits on working memory and the result was described in Fig. 3.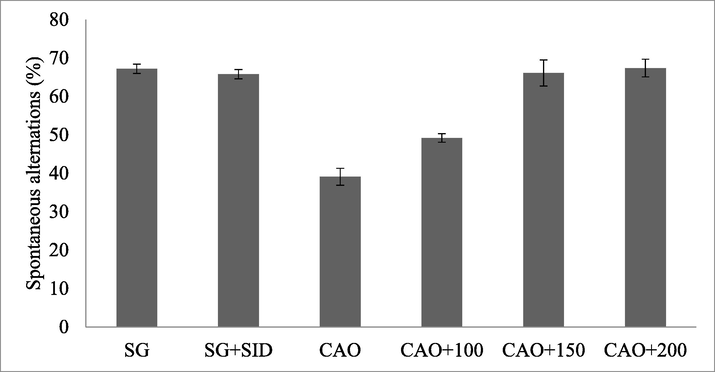
Polyphenolic compound (PP) treated ischemia-induced Albino rat and memory.
3.7 Myeloperoxidase (MPO) level in the cortex and hippocampus of experimental animal
Albino rat subjected to artery occlusion showed elevated level of MPO in the cortex and hippocampus, respectively. The amount of MPO was found to be high after 24 h. Ischemic Albino rate showed elevated levels of MPO in hippocampus and cortex and the PP treatment reduced MPO (Fig. 4A and B).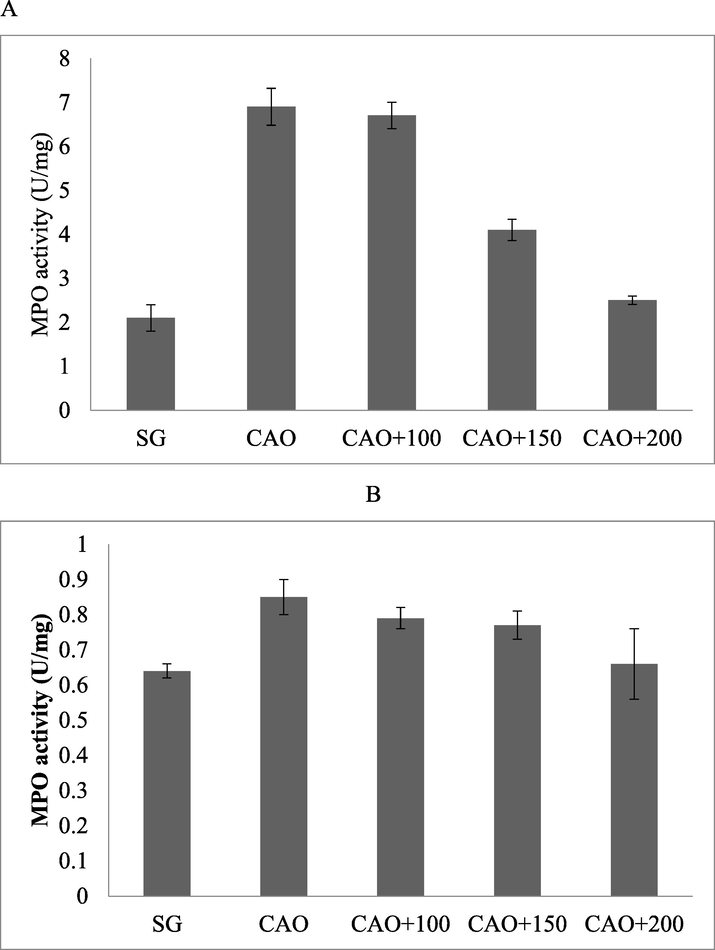
MPO activity in the cortex (A) and hippocampus (B) of mice treated with PP after artery occlusion.
3.8 Analysis of antioxidant enzymes and hydrogen peroxide
Biomarker assay was performed to evaluate the oxidative stress damage mediated by artery occlusion in the experimental animals. Enzyme assays revealed that cortex tissue is affected by occlusion mediated damage in experimental animal. Artery occlusion mediated stroke and caused reduced GPx and SOD activities than sham group of animals. Moreover, polyphenol significantly restored GPx and SOD activities. The increased concentration of polyphenol improved the biosynthesis of more antioxidant enzymes than low concentrations. In animals, reactive oxygen species are generated from the mitochondria and involved in the formation apoptotic cell death in brain injury. In our study, CAO evoked about 2.28-fold increase in ROS and further ROS generation decreased in polyphenol treated animals. Albino rats treated with different doses of polyphenol had decrease ROS amount than sham group. The present finding revealed that the neuroprotective properties of polyphenol might be associated with its antioxidant power. Caspase-3-activity level was analyzed in this study and the present research revealed that the administered PP suppressed the expression of Caspase-3-activity in experimental Albino rats (Table 2).
Experiment
Antioxidant enzymes
SOD (U/mg)
GPx (U/mg)
ROS (% sham)
Caspase-3-activity (fold)
SG
189 ± 12.5
96.4 ± 4.8
100 ± 0
1 ± 0
CAO
84.2 ± 10.5
29.1 ± 0.42
228 ± 19.2
2.1 ± 0.2
CAO + 100
141.2 ± 2.7
74.5 ± 5.8
198.2 ± 2.8
1.7 ± 0.3
CAO + 150
152.8 ± 7.4
76.3 ± 2.2
120 ± 19.4
1.4 ± 0.2
CAO + 200
162.2 ± 4.3
79.6 ± 1.1
115 ± 12.3
1.12 ± 0.12
3.9 Role of PP on cytochrome C release and caspase-3 activation in vivo trials
Polyphenols inhibited the activation of caspase-3 enzyme activity and suppressed cytochrome C release in experimental animals. It was revealed that in ischemia induced experimental Albino rats mitochondrial functions were affected and the release of cytochrome C was increased. The elevated level of cytochrome C revealed mitochondrial damage in stroke induced control Albino rat. The mitochondrial samples analyzed after 24 h of stroke increased protein expression in vehicle-treated samples than PP treatment. PP administration prevented cytochrome C release and observed reduced cytochrome C expression in ischemic cases. The 200 mg pp-treated group showed reduced expression of cytochrome C than 100 mg and 150 mg/kg pp concentrations, respectively. The administered PP suppressed the activation of caspase-3 enzyme after 24 h reperfusion and the result was described in Fig. 5.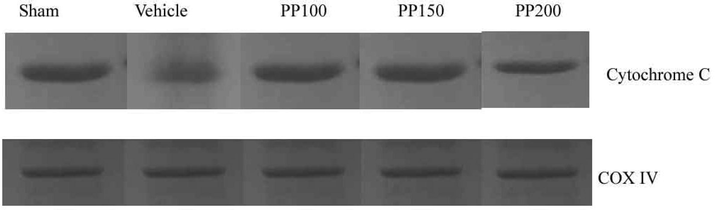
Role of PP on ischemic stroke induced Albino rat after 24 h reperfusion. Immunoblots of COXIV and cytochrome C from mitochondria sample prepared from Albino rats.
3.10 Analysis of NF-jB activation in ischemia
NF-jB activation was observed during cerebral ischemia and determined in western blot. The administered PP showed anti-oxidative stress and showed neuroprotective effect. The expression level of NF-jB was studied after 24 h of ischemia. After 24 h of reperfusion on Albino rat, upregulation of total protein sub units (p65 and phospho-p65) were determined and this upregulation process indicated expression of NF-jB sub units. Albino rats treated with PP at 150 and 200 mg/kg dosages inhibited the activation of phospho-p65 sub unit (Fig. 6).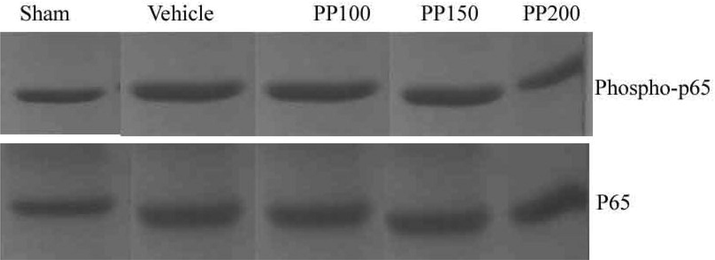
Upregulation of NF-jB subunits (p65 and phospho-p65) after 24 h reperfusion and the inhibitory activity of PP in Albino rat. Activation of p65 was observed after 24 h reperfusion treatment.
4 Discussion
The stroke preventive properties of secondary metabolites from natural sources have been reported for their anti-inflammatory, antioxidant, property against b-amyloid-induced neurotoxicity and these properties may be helpful in the prevention and treatment of cerebral ischemia. There is increasing evidence that ROS play critical role in the development of ischemia/reperfusion mediated oxidative stress in brain of experimental animals. It has been previously reported that reperfusion after ischemic stroke causes increased generation of ROS in mitochondria, and largely consumption of various endogenous antioxidants by these available radicals may lead to the increase in intracellular ROS, and these react with cellular macromolecules such as, proteins, lipids, nucleic acids, leading to severe damage of the neurons (Rychter et al., 2022). Hence, a search of molecules continues for the application of ROS scavengers and the stimulation of endogenous antioxidant molecules, for the prevention and treatment of stroke.
In recent years, an increasing interest has been shown in endophytes extract because of its safety and anti-stroke properties. The ability of phenolic compounds to cross blood–brain barrier and neuroprotective property without any side effect was reported previously (Dantas et al., 2022). In this study, we analyzed the impact of phenolic compounds in a rat model of cerebral ischemia injury and we observed that pretreatment of Albino rat with phenolic compound at 10–200 mg/kg/day improved protection against oxidative stress. The experimental animals administered with phenolic compounds improved neurological deficit. Fungal phenolic compounds inhibited ischemic neuron damage and prevented induced oxidative stress through inhibition of mitochondria-mediated apoptosis pathway. The present results revealed the therapeutic potential of fungal metabolites in ischemic stroke.
In brain, at the time of ischemic/reperfusion endogenous antioxidant enzyme activity is very important and analysis of these enzyme activities after reperfusion process is useful to check the severity of the brain cells. In our study, we analyzed antioxidant enzymes of brain muscle, and lipid peroxidation activity. Phenolic compounds was effective in inhibiting lipid peroxidation, the strong accumulation of generated free radicals due to scavenging power, and stimulating the production of GPx and SOD. The present finding is in good agreement with previous results in cerebral ischemia rat model (Abbas et al., 2022). Mitochondria are considered as the major power centre of ROS generation during reperfusion process. The elevated biosynthesis of ROS involved in the development of necrotic and apoptotic cell death by releasing various apoptogenic factors such as, caspases, apoptosis inducing factor and cytochrome C (Sims and Muyderman, 2010). Our study revealed increased release of cytochrome C from mitochondria to cytosol after 24 h reperfusion and this effect was alleviated by pretreated phenolic compounds. In our study, caspase-3 activity was found to be high after 24 h of reperfusion process, which showed increased DNA and protein damage. Moreover, phenolic compounds attenuated enzyme activity and controlled neuron apoptosis. It is observed that phenolic compound showed inhibition of ROS levels within 24 h after ischemia, however it showed inhibition of the release of cytochrome C and suppression of caspase-3 activation for 48 h. The present finding revealed that the administered antioxidant molecule delayed or blocks the expression of mitochondria-mediated aposptosis in experimental animal and reduced brain damage. Recent studies revealed that various signalling pathways involved in the generation of ROS involved in cellular damages and subsequent death in reperfusion and cerebral ischemia (Liang et al., 2008).
In this study, the neuroprotective property of polyphenol on memory deficits induced Albino rat was analyzed. The administered polyphenol suppressed the astroglial activation and inflammatory response and protected memory deficits in experimental animal through neurogenesis induction. It has been reported that polyphenols protect against neurodegenerative disorders induced by cerebral ischemia (Simonyi et al., 2005). Polyphenols protected cerebral cortex and the recovery process was associated with dose changes. The present study revealed that polyphenols improved recognition, working and special memory in experimental animal. Y-maze test was used to analyze ischemia lesions in hippocampus and striatum involved in working memory deficits in rats (Blasi et al., 2014). In the present study, spatial memory deficits were observed and the administered polyphenols attenuated these memory deficits. Polyphenols protected recognition and working memory loss in stroke induced Albino rats and improves cognitive performances
Polyphenols have neuroprotective properties and reduce brain damage and prevent memory loss in cerebral ischemia in Albino rats. Polyphenols have anti-inflammatory and antioxidant effects and reported neuroprotective properties in cerebral ischemia cases (Janarny et al., 2022). Polyphenols involved in the suppression of lipid peroxidation and inhibited cyclooxygenase and lipoxygenase activity. Myeloperoxidase activity was studied to analyze the neuroprotective property of polyphenol on memory defects caused by induced cerebral ischemia in experimental animal. Myeloperoxidase activity was elevated in ischemia/reperfusion experimental animal. Ischemia/reperfusion induced experimental animal showed neuroprotective properties in Albino rats.
5 Conclusions
The present research suggests that polyphenol exhibits potent neuroprotective properties and is useful to treat and protect cerebral reperfusion injury. The endophyte derived polypheol has neuroprotective properties in Albino rats. The ischemic Albino rats treated with polyphenol from endophyte restored memory loss and were dose dependent. The increased concentration of polyphenol stimulated the production of superoxide dismutase and glutathione peroxidase. Polyphenols reduced reactive oxygen species and protects the formation apoptotic cell death in brain and decrease levels of malondialdehyde.
Funding
None.
Acknowledgements
The authors would to thank the Sheikh Abdullah Twaijri Stroke Chair at Majmaah University, Majmaah, 11952, Saudi Arabia for supporting this work under project 3/2022.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Bioactive compounds, antioxidant, anti-inflammatory, anti-cancer, and toxicity assessment of tribulus terrestris—in vitro and in vivo studies. Antioxidants. 2022;11(6):1160.
- [Google Scholar]
- Insecticidal, antimicrobial and antioxidant activities of essential oil from Lavandula latifolia L. and its deterrent effects on Euphoria leucographa. Ind. Crops Prod.. 2021;170:113740
- [CrossRef] [Google Scholar]
- Probiotic and antioxidant potential of lactobacillus reuteri LR12 and Lactobacillus lactis LL10 isolated from pineapple puree and quality analysis of pineapple-flavored goat milk yoghurt during storage. Microorganisms. 2020;8:1-15.
- [CrossRef] [Google Scholar]
- Exploiting the potential of moringa oleifera Oil/Polyvinyl chloride polymeric bionanocomposite film enriched with silver nanoparticles for antimicrobial activity. Int. J. Polym. Sci.. 2019;2019:1-11.
- [Google Scholar]
- Medicinal plants, phytochemicals, and herbs to combat viral pathogens including SARS-CoV-2. Molecules. 2021;26(6):1775.
- [Google Scholar]
- Essential oils of two medicinal plants and protective properties of jack fruits against the spoilage bacteria and fungi. Ind. Crops Prod.. 2020;147:112239
- [CrossRef] [Google Scholar]
- Paecilomyces formosus MD12, a Biocontrol Agent to Treat Meloidogyne incognita on Brinjal in Green House. J. Fungi. 2021;2021(7):632.
- [CrossRef] [Google Scholar]
- Recognition memory impairments after subcortical white matter stroke in mice. Stroke. 2014;45:1468-1473.
- [CrossRef] [Google Scholar]
- Secondary metabolites from the endophytic fungus annulohypoxylon stygium BCRC 34024. Chem. Nat. Compd.. 2014;50:237-241.
- [CrossRef] [Google Scholar]
- Dantas, S.B.S., Moraes, G.K.A., Araujo, A.R., Chapla, V.M., 2022. Phenolic compounds and bioactive extract produced by endophytic fungus Coriolopsis rigida. https://doi.org/10.1080/14786419.2022.2115492.
- Isolation and characterization of endophytic fungi from Camptotheca acuminata. World J. Microbiol. Biotechnol.. 2013;29:1831-1838.
- [CrossRef] [Google Scholar]
- Interaction of cardiovascular risk factors with myocardial ischemia/reperfusion injury, preconditioning, and postconditioning. Pharmacol. Rev.. 2007;59:418-458.
- [CrossRef] [Google Scholar]
- Biodiversity of endophytic fungi associated with 29 traditional Chinese medicinal plants. Fungal Divers.. 2008;33:61-75.
- [Google Scholar]
- Extensive degradation and low bioavailability of orally consumed corn miRNAs in mice. Nutrients. 2018;10(2):215.
- [Google Scholar]
- Evolving therapies for myocardial ischemia/reperfusion injury. J. Am. Coll. Cardiol.. 2015;65:1454-1471.
- [CrossRef] [Google Scholar]
- Digestive recovery of polyphenols, antioxidant activity, and anti-inflammatory activity of selected edible flowers from the family Fabaceae. J. Food Biochem.. 2022;46:e14052.
- [Google Scholar]
- Genistein attenuates oxidative stress and neuronal damage following transient global cerebral ischemia in rat hippocampus. Neurosci. Lett.. 2008;438:116-120.
- [CrossRef] [Google Scholar]
- The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J Biol Chem.. 1972 May 25;247(10):3170-3175. PMID: 4623845
- [Google Scholar]
- Antioxidant effects of vitamin E and risk of cardiovascular disease in women with obesity – A narrative review. Clin. Nutr.. 2022;41:1557-1565.
- [CrossRef] [Google Scholar]
- Polyphenols in cerebral ischemia. Mol. Neurobiol.. 2005;311(31):135-147.
- [CrossRef] [Google Scholar]
- Mitochondria, oxidative metabolism and cell death in stroke. Biochim. Biophys. Acta. 2010;1802:80-91.
- [CrossRef] [Google Scholar]
- Interaction of exercise training and chronic ethanol ingestion on antioxidant system of rat brain regions. J. Appl. Toxicol.. 1997;17:329-336.
- [CrossRef] [Google Scholar]
- Advances in methods for the determination of biologically relevant lipid peroxidation products. Free Radic. Res.. 2010;44:1172-1202.
- [CrossRef] [Google Scholar]
- Isolation and characterization of some phytochemicals from indian traditional plants. Biotechnol. Res. Int.. 2012;2012:1-8.
- [CrossRef] [Google Scholar]
- PCR Protocols. Elsevier; 1990. p. :315-322.
- Diversity of endophytic fungi from roots of Panax ginseng and their saponin yield capacities. Springerplus. 2013;2:1-9.
- [CrossRef] [Google Scholar]
- Identification and determination of phenolic compounds in rapeseed meals (Brassica napus L.) J. Agric. Chem. Environ.. 2015;4:14-23.
- [CrossRef] [Google Scholar]
- Secreted monocytic miR-150 enhances targeted endothelial cell migration. Mol. Cell. 2010;39(1):133-144.
- [Google Scholar]




