

Translate this page into:
Key insights to develop drought-resilient soybean: A review
⁎Corresponding author. wuzm@jxau.edu.cn (Ziming Wu)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Soybean (Glycine max L.) is one of the most important legume crops that provide high protein contents compared to any other crop. Drought stress is one of the most devastating abiotic factors that pose severe threats to crop growth and production. Soybean growth and production are hindered by drought stress which is continuously declining yield and quality. Several studies have been conducted to unfold the genetic mechanism of drought tolerance in soybean, but to our knowledge, genetic control of drought-tolerant traits is not yet fully understood. The need for time is to make a fruitful effort to develop drought-tolerant soybean varieties that can ensure sustainability in terms of production and quality. Different soybean traits contribute to drought tolerance, and studies are being conducted to evaluate the soybean under water deficit conditions. Molecular techniques including genetic engineering, identifying quantitative trait loci (QTL), clustered regularly interspaced short palindromic repeats (CRISPR), and transcriptomes have been extensively used to develop drought-tolerant soybean cultivars. The CRISPR/Cas9 is one of the novel and most potent gene editing tools which is revolutionizing agriculture sector by targeted editing of the desired gene. Soybean offers an excellent opportunity to apply CRISPR/Cas9 because of its complex genetic network. Conventional techniques are being used to develop drought-tolerant soybean cultivars. There is still a need to integrate the available breeding tools to identify and transfer the potent genes contributing to drought tolerance. Conventional breeding methods like hybridization, backcrossing, and pedigree are suitable for breeding novel drought-tolerant cultivars. More studies are required to investigate the soybean response to drought stress at the morphological, biochemical, and molecular levels, allowing breeders to choose a proper way of improving drought tolerance. The current review provides a detailed overview of improving drought tolerance and will help conduct further research studies to develop drought-resilient soybean cultivars.
Keywords
Soybean
Drought stress
Yield
Breeding approaches

1 Introduction
Abiotic stresses threaten global food security, and the population is constantly rising (Mesquita et al., 2020). About 41% of world land is considered as dry land (Bao et al., 2019), and sudden climate change like drought stress increases throughout the globe (Cook et al., 2007). For instance, a 12-month phase of drought was seen from 2011 to 2016 (Winguth and Kelp, 2013), which caused a projected 7.5 billion dollars loss in the agrarian sector (Juenger, 2013). The agricultural statistics of China showed that around 70 billion kg of food is dumped every year because of the dry climate (Liu et al., 2013). Drought is a severe abiotic factor, adversely disturbing global crop yield and responsible for a massive disturbance in the food supply chain (Seleiman et al., 2021). By examining the current climate change pattern, it can be predicted that drought stress may become a more severe and devastating factor (Zia et al., 2021). The soybean, a wonder crop first identified in China, was nurtured about 5000 years ago and later extended to the United States of America (USA) and Europe (Zhao and Gai, 2004). The USA, Brazil, Argentina, and China are top soybean producing countries. These countries have large soybean diversity and many improved species and cultivars that showed considerable variation in yield, quality, and resistance to biotic and abiotic stresses. Wild relatives of soybean exist throughout these regions. Due to a long period of domestication and improvement, there has been significant genetic variation in soybean for all traits (Tavaud‐Pirra et al., 2009). These historical resources comprise the present germplasm population and a gene pool for the latest soybean cultivation (Czembor et al., 2021). Global soybean production during 2021 will reach to 350.72 million metric tons (Vieira and Chen, 2021).
The soybean is a significant legume crop for humans and animals (Kuromori et al., 2022). It is a powerful supplement of essential diet constituents like protein, oil, and milk (Liu et al., 2020). Drought causes 40% yield loss (Marinho et al., 2022); however, based upon the stage and the intensity of stress, this loss can go up to 80% (Guimarães-Dias et al., 2012). The primary and visible effects of drought can be seen on soybean seedlings (Han et al., 2022). Drought stress badly affects the soybean germination stage and all other growth phases (Igiehon et al., 2021). Drought stress causes severe loss in seed development at the reproductive stage. Numerous investigators have noticed the negative influence of drought on soybean seedlings and other common crops like beans (Marinho et al., 2022) and sesame crops (Boureima et al., 2011). The soybean susceptibility to water deficit conditions is linked to its growth stage. The short and moderate water shortage conditions in soybean usually do not affect productivity and quality (Vaghar et al., 2020). The soybean crop is more vulnerable to drought episodes during the developmental stage (Zhang et al., 2022b). During the seedling stage, drought stress boosts the rooting system, which is promising for soybean development, but this can enhance the water needed for pod filling (Zhengbin et al., 2011). Episodes of drought at the flowering time severely decreased the final harvest. To attain higher and steady soybean yield, breeders have categorized environmental features of drought stress to develop novel soybean genotypes with wide acclimatization against drought conditions (Jogawat et al., 2021).
China is situated in the southeastern part of the Eurasian continent, and it is one of the biggest importers of the US soybean. China observes an uneven monsoon climate and precipitation in space and time. Furthermore, the inequality of water and land assets, low-level growth, and crumbly ecological environment have revealed that China is facing repeated drought episodes (Wu et al., 2020). From 2006-to 2018, the total drought-affected area reached to 16,806.02 thousand hectares, and an estimated 21.918 billion kilograms of food was lost (Cui et al., 2021). Today, drought stress has become one of the problematic issues for China rapidly growing economy, social development, and food security (Li et al., 2020).
Unfortunately, a little piece of evidence is present on the genetic control of soybean drought tolerance (Aleem et al., 2021). As irrigation is expensive to handle the big soybean fields, irrigation is not an economical method. Hence, drought-tolerant varieties, mainly those that can germinate at low water levels, are considered one of the most feasible options (Zhang et al., 2022b). Soybean has greater tolerance against drought stress (Marinho et al., 2022) and has varied differences in changing environments. Drought tolerant cultivars have a high rate of adoptability than the cultivars grown in normal conditions. There should be a significant difference between susceptible and tolerant cultivars (Kim et al., 2022). Due to the complication of plant reactions to drought, the hunt for drought-tolerant genotypes has been a hard job, so phenotyping and documentation of traits backing to select the best genotypes under water shortage circumstances signify the main encounters in this route. Thus, a broad categorization of soybean plants in drought circumstances may designate characters backing to greater harvest constancy, thus facilitating the assortment of tolerant soybean genotypes (Giordani et al., 2019).
The fundamental stage for using available genetic material in plant breeding plans is to discover the hereditary composition of progeny, comprising the QTL with their matching alleles (Wang et al., 2020b). Documentation of the genomic areas related to drought tolerance can speed up the research progress and soybean cultivar development (Kunert and Vorster, 2020). The physiological factors, such as relative electric conductivity of leave (RECL) and relative water contents (RWC), are applicable for drought selection (Ma et al., 2004). The actual drought tolerance mechanisms are still unknown; hence use of factors like conventional breeding methods, QTL, genes, morphological characteristics, and novel techniques contributing to drought tolerance is necessary to find out a novel way of improving DT in soybean. This review discusses the latest research studies enhancing drought tolerance in soybean. The main aim was to present an updated picture of different techniques to improve drought tolerance in soybean.
2 Effect of drought stress on soybean
Drought episodes significantly disturb soybean yield and quality (Fig. 1) and cause severe danger to global food safety (Xiong et al., 2021). Drought stress reduces stomatal conductance, total biomass, yield, and other essential characteristics (Basal and Szabó, 2020a) (Table 1). A decrease in chlorophyll contents and the electron transport network is also linked to drought stress-induced changes. Reactive oxygen species (ROS) cause oxidation in plant and lead to the disturbances of several processes (Imran et al., 2021), which are then accumulated in large applications (Shigeoka et al., 2002) and cause harm to genes modification, causes protein breakdown and inactivation of enzymes (Mahajan and Tuteja, 2005). ROS in higher concentrations causes injuries to the cells and affects numerous routes like soybean germination and growth (Dowling and Simmons, 2009). At the grain filling phase, drought stress decreases the soybean growth ratio by reducing photosynthetic activity and disturbing the assimilated supply chain converted to storage blends. Earlier studies found that average and drastic drought stress can reduce the seed growth by 9 to 35% (Nakagawa et al., 2018). The sucrose contents of soybean seeds reduced considerably by 9% in 7 days of drought stress (Egli and Bruening, 2004). This evidence provides novel understandings of drought stress mechanisms and genomic approaches. Severe drought stress impairs dry matter stock and photosynthetic efficacy (Cui et al., 2021). Leaf temperature rises because of the reduction in cooling effect due to the reduced transpiration activity (Igiehon et al., 2021). Water deficiency decreases the expansion of plant cells and enhances membrane permeability and causes cell death. The main changes comprising improved growth of roots and stomatal closure are related to various genes and corridors, containing signal transduction, protein breakdown, balancing of phytohormones synthesis, and carbohydrates breakdown (Fu et al., 2022). Moreover, drought also induces changes in the arrangement of fats, protein, and carbohydrate content in soybean seeds (Xiong et al., 2021). Soybean photochemical efficiency was reduced by exposure to drought stress (Basal et al., 2020). In the same way, drought stress caused a severe decline in grain yield (Chandra et al., 2021).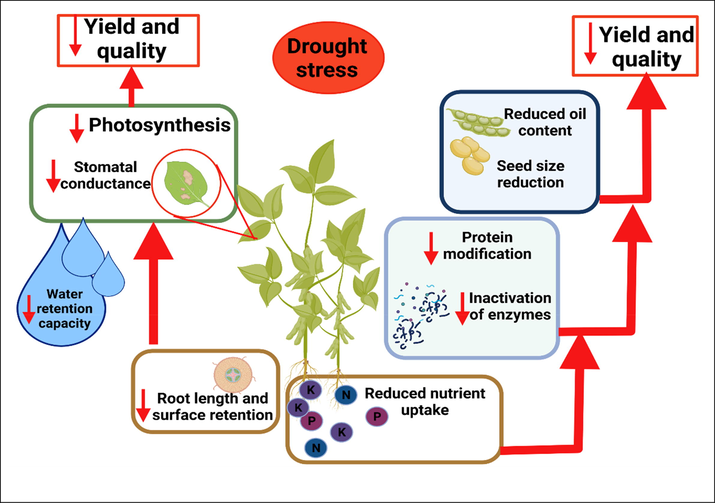
Drought stress leads to several harmful effects on soybean crops like reduction in photosynthesis rate, nutrients uptake, protein modification, reduction in oil contents. Drought stress reduces root length and also leads to the inactivation of enzymes.
Effect of drought stress
References
Drought stress affected relative water contents, and fresh root weight
(Waqar et al., 2022)
Drought stress reduced the number of fertile pods in soybean
(Mejaya et al., 2022)
Drought stress reduced chlorophyll contents
(Sadeghi et al., 2021)
Drought stress caused the stomatal closure after soybean exposure to severe stress
(Seleiman et al., 2021)
Drought stress reduced the grain yield of soybean
(Chandra et al., 2021)
Reduced sugar metabolism, nitrogen, and soluble protein contents
(Du et al., 2020)
Photochemical efficiency reduced under severe drought stress
(Basal et al., 2020)
Reduced cadmium update
(Bashir et al., 2019)
Drought stress decreased nutrients storage reserve
(Wijewardana et al., 2019)
DS decreased the oil contents of soybean at the grains filling stage (Fig. 1). Drought stress considerably reduced the function of genes responsible for lipid synthesis, pyruvate kinase, and transporter proteins synthesis in developed seeds between the 19 and 29 days of severe drought (Nakagawa et al., 2018). Extreme drought stress decreases the soybean seed dry weight by reducing the lipid content in seeds, decreasing their synthesis, and stimulating their degeneracy (Wei et al., 2015). More studies are being conducted on lipids regulation and breakdown of protein under drought stress. In soybean, drought stress reduces the nitrogen concentration by reducing nitrogen metabolism efficiency, which involves a reduction in enzymatic activity in nitrogen metabolism (Du et al., 2020) (Table 1). Water scarcity also affects the availability of phosphorous. An experiment showed that drought stress limited the accumulation and translocation of phosphorous (P) in soybean seeds (Fig. 1) (Jin et al., 2006). Drought stress interrupts the plant water relation in susceptible cultivars. RWC and leaf water potential were reduced under drought stress (Chowdhury et al., 2017). Water retention capacity was also studied in a drought-prone environment. Hence, more studies are required to study the effects of drought stress on various growth stages. Soybean should be exposed to different levels of drought stress for a comprehensive understanding of the mechanism of drought stress and changes that occur after stress.
3 Challenges of drought breeding
Breeding for drought stress or water deficit conditions is difficult (Marinho et al., 2022) mainly due to the unavailability of data about the genomic regions for drought tolerance. For this reason, it may be problematic to develop a genetic marker for soybean reaction to drought stress (Mangena, 2020). A marker is a difference in a gene order on the chromosome and its relationship with a typical feature (Younis et al., 2020). After this, tolerant plants are identified via marker-assisted selection (MAS) (Sreenivasa et al., 2020). The intensity and duration of drought stress may vary (Wang et al., 2020a), which puts an obstacle in assessing new varieties. Hence, plant researchers usually collect data from different locations and examine the differential reaction of the soybean to drought stress across changing areas (Kannenberg et al., 2021). Improving drought tolerance will not result in stable yield at many locations, making it hard to merge values across areas (Feng et al., 2020). Drought resilient cultivars perform well under normal circumstances. It is not clear that yield under typical situations should be enhanced besides rise in drought episodes (Zhou et al., 2021). The proposals showed that drought stress causes substantial yield loss, and therefore, it is significant to choose cultivars with higher drought tolerance (Zhou et al., 2021). Though numerous reviews on the consequences of drought stress in grain legumes are accessible, a complete review on the challenges of drought stress in soybean to improve its output is not presented (Farooq et al., 2017). Hence, challenges of drought breeding in soybean should be solved by use of latest breeding tools which will unfold the genetic mechanism of drought tolerance.
4 Mechanisms of drought tolerance
The mechanism of drought stress tolerance is advantageous to farmers to adopt as a conventional method to escape the drought period (Moloi and van der Merwe, 2021). In this process, plants rapidly complete their life cycles to avoid drought stress (Moloi and van der Merwe, 2021). The escape mechanism in soybean is mainly applied in the US and cultivar is identified as early growing genotype (Puspitasari and Arwin, 2021). The soybeans are planted in March and complete their lifecycle in early July when drought stress is at the peak (Manavalan et al., 2009). Early soybean planting results in rapid growth, extensive canopy development, and high yield (Iqbal et al., 2019). Therefore, plant breeders can assist the farmers in adopting a drought escape system by developing cultivars best suited to this system. Early flowering is an essential feature of drought tolerance mechanism, and it has a significant effect on yield when plants go under severe drought stress (Chandra et al., 2021). The early flowering trait is developed by classical breeding plan by making noticeable selection in the F2 population for plants with early flowering character (Su et al., 2021). The plant has a waxy layer and leaf abscission mechanisms to counter drought stress (Rasheed et al., 2020b) (Fig. 2). Soybean response to drought stress at morphological, physiological, and molecular levels needs more studies to identify the potent factors that help soybean cope with severe drought stress. Future studies should focus on developing early maturing soybean cultivars which can escape drought to maintain growth and yield.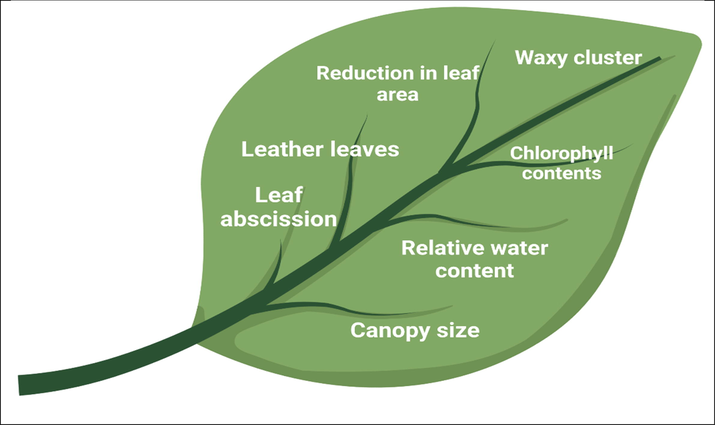
A graphical display of soybean defense response against drought pressure. Drought causes activation of several responses in soybean like waxy cluster, canopy size, leaf abscission, reducing leaf era, and maintaining chlorophyll.
5 Conventional methods of improving drought tolerance
The drought-tolerant soybean varieties are naturally selected based on phenotypic differences among the plants grown in drought circumstances. There are lot of traditional breeding methods like, introduction, mass selection, and hybridization (Fig. 3) used to develop drought tolerant cultivars. Hundreds of drought tolerant cultivars of soybean have been developed and released by using these methods (Pathan et al., 2007). Classical breeding techniques for developing drought-tolerant soybean genotypes are time-consuming, and breeders have specific difficulties because drought tolerance is a quantitatively inherited trait (Charlson et al., 2009). Therefore, if quantitative genetics can recognize QTL linked to drought tolerance, breeders would have a more helpful direction to quickly develop better genotypes having improved traits under drought situations, for instance, via MAS selection (Liu et al., 2020). Hence, drought-tolerant plants conventionally use a drought escape mechanism.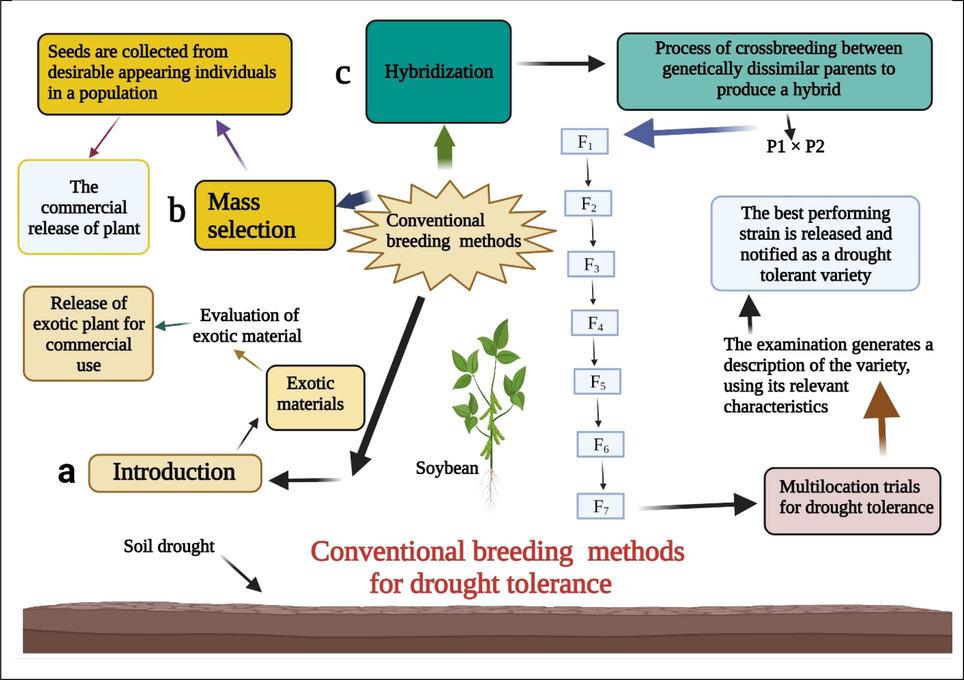
The depiction of several conventional breeding techniques leads to the breeding of drought-tolerant genotypes in soybean. Plant breeding approaches like introduction, hybridization, and mass selection have been broadly used. Modifying these methods would promise to improve soybean growth and yield under drought stress.
5.1 Use of agronomic techniques
Use of fertilizers has been broadly practiced in crop production. Nonetheless, quantity of fertilizers should be measured, as too much amount adversely affects plants. Protein quantity of soybean has been improved by adequate irrigation and N-fertilizer rate. At the same time, oil quantity was reduced by water application and by a higher dose of fertilization under drought stress (Basal and Szabó, 2020b). The use of phosphorous effectively improved the yield and quality traits of soybean. A study was performed to assess the consequence of P fertilizer on soybean root growth under drought conditions. According to the results P treated soybean genotypes showed better root growth, and it is proven that adequate fertilization is a critical need to mitigate the adverse impacts of drought (Hansel et al., 2017). The combined effect of zinc (Zn) and manganese (Mn) improved the soybean yield and quality under drought conditions. The soybean cultivars like Williams and Zan were grown under drought conditions. The proline and linoleic acid contents were improved considerably, and the entire contents of protein and linolenic acid were higher in cultivar (Williams). Consequences of zinc were noticed in root length and linoleic acid; hence the maximum leaf proline was achieved using manganese sulfate fertilizer (Zolfaghari Gheshlaghi et al., 2019). Likewise, the consequences of P were studied on the seedlings and physiology of soybean under water deficit conditions. P improved soybean root and growth traits and biochemical attributes (Abd El–Mageed et al., 2017). Plant hormones have a crucial function in soybean growth under drought stress (Imran et al., 2021). Melatonin (MT) is a bioactive indolamine hormone that controls various physiological procedures during development and increases tolerance against drought stress.
Treatment of soybean plants with melatonin (Fig. 4) at a concentration of 50 and 100 µM considerably alleviated the harmful consequences of drought on chlorophyll content (Imran et al., 2021). The ethylene triggers a defense response in seedlings and other reactions to numerous stresses. Numerous researchers studied the influence of ethylene on plants under drought. A higher ethylene concentration in soybean improved the growth and production under drought circumstances. Hence, combining this evidence showed ethylene synthesis and signaling network contribution to drought tolerance (Arraes et al., 2015). Cytokinin (CK) response and movement are regulated by signaling and related to stress incentives to form molecular and biochemical reactions that endorse the reworking to ecological fluctuations, containing source and sink alteration (Peleg et al., 2011) and grain yield (Kambhampati et al., 2017). This evidence showed the relationship of CK with other hormones, abscisic acid (ABA), and salicylic acid (SA) (Salek Mearaji et al., 2021), regulating drought stress response in soybean. The consequence of SA and ascorbic acid was studied on soybean morphology and yield traits under drought stress. The treatments include ascorbic acid (1000 mg/1L) and T4- salicylic acid (10 ml/1L). SA improved grain weight (Fig. 4), and ascorbic acid improved soybean drought tolerance and growth (Alimento et al., 2021). Studies recommend that foliar use of SA could alleviate drought pressure, as demonstrated by the yield traits of soybean crops (Alimento et al., 2021).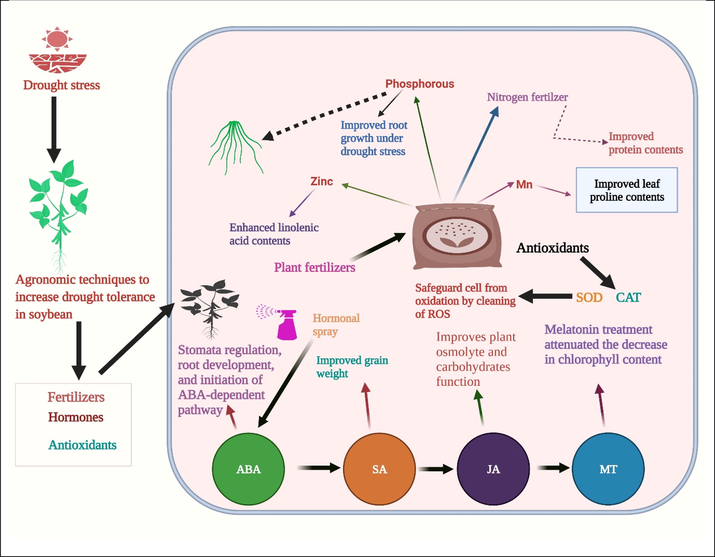
Role of agronomic techniques to improve tolerance against drought stress in soybean. It involves use of fertilizers, hormones and antioxidants which safeguard soybean plant from harmful effects of drought stress.
Jasmonic acid (JA) improves plant osmolyte and carbohydrates against drought conditions. The primary function of growth hormones in improving photosynthesis in drought conditions has been studied. Furthermore, JA can alter the synthesis of metabolites in soybean (Anjum et al., 2011). Mohamed et al. (2017) studied the consequence of JA on the water and oil contents of soybean under drought stress. They found a substantial rise in fractionation of cell wall under stress conditions (Mohamed et al., 2017). In another experiment, effect of JA at the concentration of 100 µM was studied on protein contents in soybean cultivar (Giza 22) under drought stress. Results revealed that JA reduced the consequences of drought stress on soybean (Hassanein et al., 2009). Hence, it is determined that using a high level of hormones under drought circumstances increases soybean growth and development. Foliar application of hormones would be critical to mitigate the adverse effects of drought stress.
5.2 Antioxidant defense system
The critical role of antioxidants is to protect the plants from the damaging impacts of ROS (Rahman et al., 2021). Under drought situations, antioxidants like, superoxide dismutase (SOD) and catalase (CAT) usually surge, and reduce the concentration of malondialdehyde (MDA) to improve the soybean growth (Wu et al., 2019a; Wu et al., 2019b) (Fig. 4). The increased concentration of antioxidants in soybean enhanced growth, like a rise in plant height, enhanced leaf growth, and improved the accumulation of dry matter stock (Wu et al., 2019a; Wu et al., 2019b). Through this development, variations in the proportions of diverse endogenous hormone concentrations can result in different physiological factors that control the growth of tissues. The increased activity of antioxidants could efficiently alleviate membrane injury and lipid peroxidation. In addition, soluble sugars and proline contents also increase (Luo et al., 2014) and eventually cause a growth compensation (Imran et al., 2021). The role of antioxidants is critical to mitigate the adverse effects of drought stress in soybean. Their role should be studied by exposing plants to varied levels of drought stress. These antioxidants enzymes would help to sustain soybean growth and yield by detoxifying the ROS and prevent oxidation in the cell.
6 Molecular tools toward drought-resilient soybean
Molecular techniques are widely used to bring drought tolerance in soybean (de Freitas et al., 2022). A gene AtMYB44 from Arabidopsis thaliana was successfully transferred using the Agrobacterium-mediated gene transfer method into soybean genotype, which enhanced yield and quality under drought conditions (Seo et al., 2012). The optimization of the Agrobacterium-mediated gene transfer technique and regeneration of soybean cultivars using the tissue culture technique will assist in the successful engineering of soybean for drought stress tolerance (Raza et al., 2020). Remarkably, genetic manipulation practices are being used to develop soybeans grown in laboratories and glasshouses and accepted for commercial cultivation. This review has compiled all recent studies on soybean drought tolerance using molecular approaches.
6.1 Quantitative trait loci (QTL) to improve drought tolerance
In recent studies, at least one thousand QTL signifying more than hundreds of agronomically essential characters have been detected in soybeans (Grant et al., 2010). Present evidence on all identified QTL in soybean is accessible on: https://soybase.org. Although numerous QTL are documented in the soybean, the pyramiding of QTL controlling the same trait is difficult in breeding plans. Due to soaring needs, breeders are using molecular approaches to develop the drought-tolerant cultivars of soybean. Meanwhile, molecular markers recognized genetic alternatives for diverse drought-associated characters with higher accuracy. Molecular markers are vital for developing genetic linkages maps, gene diagramming, and evaluating genetic resources (Satpute et al., 2020).
The mechanism of DT in soybean is highly complicated and is a consequence of complex systems of numerous genes. Breeding for drought tolerance has been recognized as a significant objective for soybean breeding. Multiple investigations have shown that genetic studies predicted noticeable differences for given traits under drought stress circumstances (Giordani et al., 2019). Identifying QTL for soybean drought tolerance is critical to ensure plant growth under drought stress. QTL for yield under drought conditions can be transferred into drought susceptible cultivars to enhance productivity. Yield is the most affected trait under drought stress. Identifying QTL maintaining higher yield under drought stress would be a promising approach to ensure yield stability in stress-prone environments. In an experiment, Lee et al. (2021) identified two yield-related QTL on chromosomes 13 and 17 under rain-fed conditions. The line with PI 471,938 allele yielded higher than lines with Hutcheson alleles. Wang et al. (2020b) identified several QTL likely to link with DT using an efficient restricted two-stage multi-locus multi-allele genome-wide association study. The identified QTL conferred drought tolerance in soybean cultivars. Therefore, this study evidenced the complexity of DT genetics and its improvement at the molecular level. In another experiment, 10 QTL contributing about 12.9% variation were recognized on chromosomes 2, 7, 10, 14 and 20. These QTL and associated markers could be applied in MAS selection to speed up the drought tolerance breeding in soybean (Dhungana et al., 2021).
In an experiment, a drought sensitive (Zhonghuang 35) and tolerant variety (Jindou 21) genotypes were crossed to obtain F6:9 recombinant inbred lines (RILs) population. The traits like planting height and seed weight were selected as drought tolerance indexes. A total of 23 QTL for DT were identified. Among the identified QTL, two new major QTL, qSWPP19, and qSWPP17, were recognized for seed weight per plant (Ren et al., 2020) (Table 2). Zhang et al. (2021) constructed a linkage map using single nucleotides polymorphism markers (SNP) and detected 18 QTL positioned on seven soybean chromosomes. The QTL analysis revealed that two QTL, qPH7-4, and qPH7-6, were major DT QTL. These results laid the basis for MAS to improve drought tolerance in soybean seedlings. The identification of DT at the seedling stage can be highly considered to unfold the complex mechanism of DT in soybean. The major DT QTL, qRL11, was identified in a study and strongly contributed to DT in soybean (Liu et al., 2020). The soybean yield is at constant risk of decline under the increasing threat of drought. Canopy temperature is an indirect measurement of stomatal conductance, and it may help differentiate the genotype response towards drought stress. In a study conducted by Bazzer and Purcell (2020), 11 significant QTL positioned on eight chromosomes were identified, independently describing the phenotypic difference between 4.6% and 12.3%. The QTL, qCT3, was a potent QTL for drought tolerance. In another study, Khan et al. (2018) used the RILs population to conduct a QTL mapping analysis for drought tolerance in soybean. They have identified a novel QTL, qRSL1.1 for shoot length, which positively contributed to the drought tolerance in soybean. Hence shoot length (SL) can also be used as a drought tolerance parameter for selection of drought tolerant genotypes of soybean.
QTL
Chromosome
Trait
References
qRDW1
1
Root dry weight
(Gai et al., 2007)
qSatt255
5
Drought tolerance
(Zhang et al., 2012)
qSW-Gm02
2
Slow wilting canopy
(Abdel-Haleem et al., 2012)
qRWC-M−2, qCSS-G5
2, 5
Relative water content, and concentration of soluble sugar
(Yang et al., 2014)
qRSL1.1
1
Shoot length
(Khan et al., 2018)
qRPL2-1
2
Relative plant height
(Khan et al., 2019)
qMPH1.1
1
Plant height
(Wang et al., 2020b)
SWPP17, SWPP19
17, 19
Seed weight per plant
(Ren et al., 2020)
qRL11
11
Root length
(Liu et al., 2020)
qCT3
3
Canopy temperature
(Bazzer and Purcell, 2020)
qPH7-4
7
Plant height
(Zhang et al., 2021)
qYLD3
3
Yield
(Lee et al., 2021)
qWDC7.1
7
Morphological traits
(Dhungana et al., 2021)
Previous studies used recombinant inbred line population (RILs) to detect the QTL for wilting traits under field conditions. The QTL, qSW-Gm02, was associated with canopy tolerance in soybean (Abdel-Haleem et al., 2012) (Table 2). Using these QTL for QTL pyramiding, drought tolerance can be effectively improved to maintain the soybean yield in drought-prone environments. However, additional research is needed to recognize and clone the novel regions related to drought tolerance in soybean.
6.2 QTL for slow canopy
In many studies large-scale screening has been done using soybean germplasm to identify traits linked with drought tolerance (Valliyodan et al., 2017). The latest study recognized two significant QTL, qSW_Gm06 and qSW_Gm10, associated with slow canopy (Ye et al., 2020). Genotypes with a slow canopy wilting appearance showed that PI 416,937 was least prone to drought episodes compared to the lines grown in the field conditions. A larger nodule dry weight was witnessed in plants than in drought-sensitive check cultivars (Begum et al., 2022; Jiang et al., 2022). This proposes that a slow canopy wilting nature may be attained through root systems that permit better accessibility to water channels. Due to a better root system, PI 416,937 achieved better transpiration under drought stress (Abdel-Haleem et al., 2012). Meanwhile, in the 1990 s, the slow canopy wilting phenotype was previously used as a marker to sort for drought tolerance in soybean (Charlson et al., 2009). Numerous drought-tolerant lines have been developed from these selections (Devi et al., 2014).
One encounter that breeders look at using these QTL is the complex nature of canopy wilting traits under drought circumstances. The maximum identified QTL were uneven across independent locations and populations. For example, the major QTL on chromosome 12 was documented in all five areas in Benning × PI 416,937 (Abdel-Haleem et al., 2012). Still, it was not identified in any other populations described by Hwang et al. (2015). In more progressive generations, such as BCnF2, QTL validation should confirm each separate QTL for a particular trait. It shows that assembling all validated QTL in a similar leading background by MAS is essential to shaping the drought tolerance presented in the original population. Newly, Chamarthi et al. (2021) recognized loci related to wilting canopy traits using 200 different maturity groups. In the neighborhood of 188 significant SNPs, 183 genes were identified for both corresponding SNPs and new SNPs. These genes may be linked with transpiration. The validated genomic areas may be a significant source for pyramiding favorable alleles to improve drought tolerance in soybean (Chamarthi et al., 2021). Evaluation of soybean population at seedling stage under drought stress would lead to identify the tolerant lines and can be used to develop cultivars tolerant to drought stress. Screening of tolerant lines would be useful to identify the QTL for drought tolerance in soybean population.
6.3 Use of genome editing technique towards drought tolerance in soybean
Nowadays CRISPR/Cas9 novel gene-editing technique yields promising results for successful drought tolerance in soybean (Yuan et al., 2021) (Fig. 5). Gene manipulation using CRISPR/Cas9 has been an effective technique for crop improvement. However, its effectiveness can be enhanced, particularly for soybean crops with a polygenic nature. Numerous abiotic stresses have reduced plant growth and development, limiting crop yields worldwide (Rasheed et al., 2020a; Rasheed et al., 2020b; Rasheed et al., 2021). Because of the complexities of drought stress, genome alteration for drought tolerance has only recently been demonstrated (Rasheed et al., 2020b). Drought signaling genes and overexpression of transcription factors promote the accumulation of signaling molecules and metabolites, which improves drought tolerance in plants. Conversely, tolerant genes improved the tolerance against drought, improving the antioxidant activities and maintaining the hormonal balance (Rasheed et al., 2020b). The CRISPR/Cas9 method was applied to edit new alleles of gene (OST2), a significant plasma membrane H + ATPase essential for stomatal reaction (Osakabe et al., 2016). Mutation in transgenic plants were detected with high mutation efficiency (>32 percent) and no off-target alterations using a modified CRISPR/Cas9 system with a truncated sgRNA (tru-sgRNA) and Cas9 combination. When the stomatal response was measured under ABA-induced circumstances, it was discovered that OST2 CRISPR mutants exhibited a much higher degree of stomatal closure and a lower amount of transcriptional water loss than the wild type. A CRISPR/Cas9-induced mutation at the OST2 gene improved drought tolerance by improving stomatal responsiveness.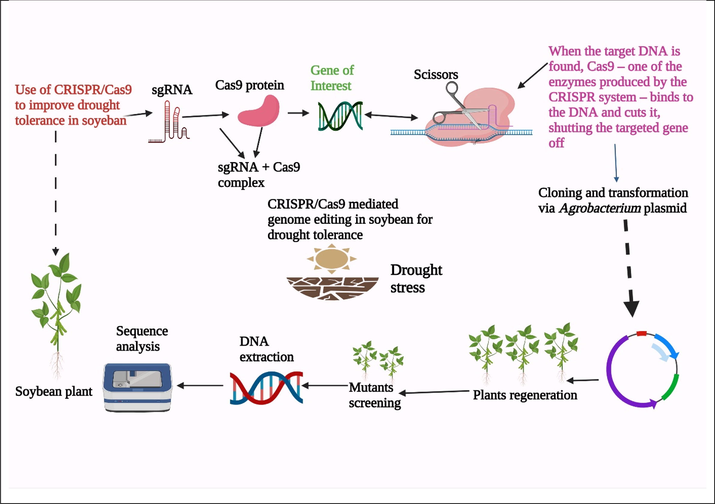
Mechanism of CRISPR/Cas9 based genome editing in soybean. It involves the complex of sgRNA and Cas9 protein, unwinding of DNA by sgRNA, cleavage of gene by Cas9, cloning of gene by a vector, transformation of genes using Agrobacterium plasmids, regeneration of plants, screening of the desired mutants, and DNA extraction to check the integration of targeted gene. In the end, sequence analysis is done to confirm the gene expression.
According to a comprehensive molecular investigation, ABA acts as a leading player in drought reaction in plants by modifying the function of stress-related genes and modulating stomatal closure to avoid (Shan et al., 2022) water loss. The bZIP family of transcription factors, also known as ABA response element-binding protein/ABRE binding factors (AREBs/ABFs), is a critical component of ABA signaling (Nakashima et al., 2014). Drought tolerance was improved when AREB1 expression was increased, but knocking out of AREB1 led to significant vulnerability to drought stress (Singh and Laxmi, 2015). AREB1 acts as a primary factor of ABA production, antioxidant signaling, and osmotic protection by regulating many genes downstream of the ABA signaling pathway (Li et al., 2013). Application of CRISPR/Cas9 will bring targeted editing of any desired gene to develop drought tolerant soybean cultivars. Use of different Cas variants as well as base and prime editing is recommended for efficient editing of gene for drought tolerance. As a result, AREB1 could be a good target for increasing drought tolerance in soybean. Roca Paixão et al. (2019) studied whether the CRISPR initiation (CRISPRa) system that edit AREB1 might play a role to increase drought stress tolerance in Arabidopsis. Arabidopsis histone acetyltransferase 1 (AtHAT1) indorses gene expression initiation by closing chromatin to a relaxed state. The plants exhibited higher chlorophyll content and quicker stomatal aperture under water scarcity, in addition to a better existence rate after drought stress.The GmNAC8 gene was confined in the nucleus, and the action of the GmNAC8 gene was considered significant in response to drought stress. Furthermore, the GmDi19-3 expression pattern exhibited the same pattern as the GmNAC8 gene against drought stress. Therefore, the above results showed that GmNAC8 works as a positive controller of drought tolerance in soybean and confirmed that GmNAC8 possibly acts by interrelating with additional constructive guiding gene-like, GmDi19-3 (Yang et al., 2020). These results offered an excellent opportunity for CRISPR/Cas9 (Fig. 5) for targeted gene editing. Hence, applying different CRISPR systems would yield promising results in developing drought-tolerant cultivars.
6.4 Genetic engineering technique and transcription factors for drought tolerance in soybean
Genome engineering methods offer feasible prospects to enhance soybean improvement (Arya et al., 2021; Lohani et al., 2020). The genes regulating characters, such as flowering time and disease resistance, have been recognized and used for soybean development (Arya et al., 2021). Likewise, high throughput genome and transcriptome sequencing have led to documentation of necessary transcriptional controllers of the soybean drought mechanism. The DREB (Nguyen et al., 2019), MYB (Chen et al., 2021a), and WRKY (Wei et al., 2019) are the prominent transcription factors families which facilitate the plant reactions under stress conditions. RNAi and CRISPR/Cas9 have provided evidence of how complex gene networking systems control tolerance against drought in soybean. For instance, Yang et al. (2020) noted that the GmNAC8 transcription factor substantially contains the tolerance against drought in soybean. GmNAC8 was replicated under the control of 35 s promoter for gene over-expression, and CRISPR/Cas9 was applied to edit the GmNAC8. Over-expression and knockdown lines were developed, and lines with over-expression of GmNAC8 showed a substantial increase in antioxidant activities and proline accumulation, which conferred the drought tolerance.
The US Department of Agriculture (USDA) recently allowed the production of soybeans that produce the HaHB4 transcription factor found in sunflowers (Helianthus annus). HaHB4 is a drought-tolerant sunflower transcription factor whose overexpression in Arabidopsis improves drought tolerance (Manavella et al., 2008). The constitutive 35S promoter and the inducible HaHB4 promoter were used to clone HaHB4. Ribichich et al. (2020) investigated soybean with HaHB4-expressing in a glasshouse and the field. When genotype to environment contact (GE) was studied, transgenic line identified as b10H did best in field experiments. Under warm and dry circumstances, the b10H soybeans exhibited a higher yield (seed number), which was not balanced by lower seed weight. Compared to WT, b10H generated 43.4% more profit under water shortage situations. In addition, b10H soybeans had larger epicotyls, internodes, and xylem diameters than WT plants. Furthermore, b10H plants showed a considerably higher photosynthetic rate (at the R5 and R6 stages). Heat shock proteins homologous to Arabidopsis thaliana HSC70-1 (At5G02500), HSFB2A (At5G62020), Hsp81.4 (At5G56000), and HOT5 (At5G43940) transcripts were found to be differently regulated in soybeans expressing HaHB4 (Ribichich et al., 2020). The line with HaHB4 has acquired regulatory approval in Argentina, Brazil, Paraguay, and Canada because of outstanding field performance (Businesswire, 2021). Several genes and TFs have been identified and transformed to develop drought tolerant soybean cultivars (Table 3, Fig. 6). These findings revealed that GE is a potent way of improving soybean drought tolerance by transforming resistant genes into susceptible cultivars.
Genes/TFs
Role
References
GmDREB2A;2 CA
Increased yield and growth traits
(Marinho et al., 2022)
GmEF8
Enhanced drought tolerance in soybean
(Zhang et al., 2022a)
GmXTH1
Improved root system, relative water content and activity of antioxidant enzymes
(Zhang et al., 2022b)
GmPOD40
Increased photosynthesis and antioxidant activities
(Aleem et al., 2022)
GmTGA15
Enhanced chlorophyll content
(Chen et al., 2021b)
LHY
Enhanced drought tolerance
(Wang et al., 2021)
GmMYB14
Decreased leaf size, leaf petiole length, leaf petiole angle and improved yield
(Chen et al., 2021a)
GmSOG1
Overexpression of gene enhanced anti-oxidation enzyme accumulation and increased drought stress tolerance
(Poku et al., 2021)
ERECTA
Enhanced greater stomata density and conductance to increase stress tolerance
(Berchembrock et al., 2021)
AtAREB1
Plants with overexpression of AtAREB1 maintained higher photosynthetic rate
(Fuhrmann-Aoyagi et al., 2021)
DREB
Improved drought stress tolerance
(Zhou et al., 2020)
GmNFYA13
Plants with GmNFYA13 showed increased transcript levels of stress-related genes exhibited ABA hypersensitivity
(Xu et al., 2020)
WRKY20
Enhanced yield, plant height, and higher seed rate
(Ning et al., 2017)
GmWRKY27
Increased proline contents
(Wang et al., 2015)
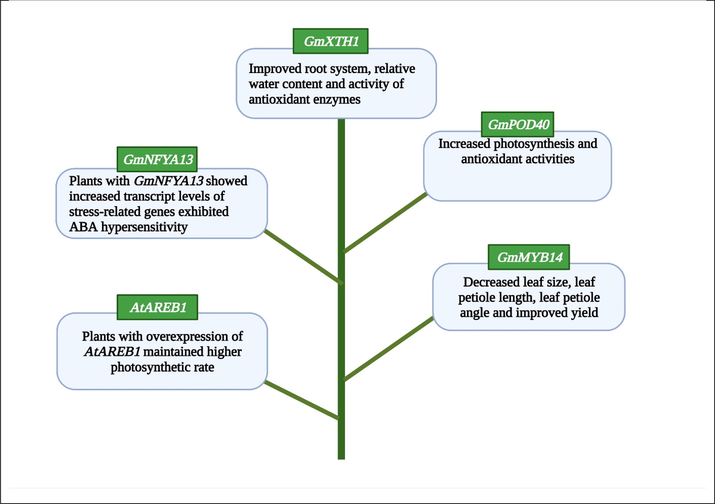
The application of genome engineering in soybean leads to development of several transgenic lines against drought stress. The identification and transformation of different genes lead to the development of drought tolerant soybean cultivars.
6.5 Use of transcriptomic and proteomics techniques
The transcriptomic analysis is essential to recognize the consequences of drought at the transcriptomic level. The RNA-Seq technique is often applied to investigate the transcriptomes involved in water scarcity in soybean. The RNA-Seq method is used to study the phases of seed development (Severin et al., 2010). Many defense systems activate numerous reactions that enhance the plant's tolerance to difficult ecological situations. Identifying stress signaling pathways and sequential molecular, physiological, and biochemical reactions triggered by the signal transduction corridor is the primary stage in the stress reaction in soybean (Le et al., 2012). In soybean, plenty of proteins are responsible for photosynthesis, plant growth, development, and stress response. Defensive mechanisms against drought stress were improved as described by the plenty of proteins accountable for redox balance, and breakdown at the vegetative phase. The protein downregulation at the flowering stage also added to nitrogen remobilization in soybean (Sharma et al., 2018).
By applying the proteomics technique, Yu et al. (2016) matched the different protein abundances of two soybean cultivars; one is drought resilient, and the other is drought sensitive. Proteins were taken and sorted using the electrophoresis method. The results showed that 38.5% of proteins are accountable for metabolic pathways and 36.4% for defense pathways. Drought stress-triggered increased proteins in different quantities (Yu et al., 2016). The proteomics technique have been developed to modify biological materials (Rai et al., 2021). The proteomics study of a superior soybean genotype (Jidou17), along with its parental genotypes to assess the outstanding traits, was done using the (iTRAQ-based) proteomic method (Qin et al., 2013).
Yahoueian et al. (2021) performed a comparison study to check the level of leaf proteome in two soybean genotypes, drought-sensitive (GN-2032), and drought resilient (GN-3074) by exposing them to drought episodes. Proteins were removed from leaves of plants from both control and stressed seedlings by applying the trichloroacetic acid–acetone precipitation method and examined by gel electrophoresis. The 26 proteins exhibited noteworthy changes in one cultivar. The recognition of 20 proteins by mass spectrometry showed a synchronized protein expression responsible for cellular breakdowns comprising photosynthetic pathways and methyl transduction, allowing plants to curb drought situations. Our outcomes recommend that the soybean plant encounter drought stress by using the stay green trend via synchronized gene action at the vegetative phase (Yahoueian et al., 2021). On the whole, about ninety-seven proteins were altered individually during the experiment. The number of proteins involved in protein production considerably improved during drought stress. The redox proteins enhanced drought tolerance. Hence, some standard proteins related to photosynthesis, and signaling network are linked with a response to drought stress in soybean (Qin et al., 2013). Particular proteins, like lipoxygenases, nitrogen, and carbon metabolism, are comparable to root proteins and were noted to be downregulated (Gil-Quintana et al., 2015). Studies revealed that soybean showed a high degree of tolerance. These studies provided the foundation for research on soybean engineering for successful yield and quality improvement under drought stress (Yahoueian et al., 2021). Identifying novel genes and proteins in soybean is possible by exposing plants to severe drought stress. The transcriptome analysis of drought-stressed plants would identify critical genes regulating drought tolerance in soybean and stress-associated pathways.
7 Conclusions and future perspective
The green revolution increased the total plant production but increasing global warming and world population are continuously challenging our ability to feed the entire world. Therefore, there has been a rise in interest in increasing crop yields and quality because of crop yield loss by severe drought stress during the last few decades. The current review aims to critically elaborate the numerous ways of improving drought tolerance and sheds light on future research directions. The response mechanism is highly studied at the morphological and molecular level in plants. Genetic variations in root development, leaf angle, and stomatal closure are linked with the morpho-physiological type of responses. Several hormones like ABA, CK, SA, and fertilizers are applied to mitigate the harmful consequences of drought episodes. The phytohormones level in crops usually enhances upon drought stimulus and ultimately activates these stress responsive pathways.
The use of hormones is increasing to mitigate the harmful impacts of drought. This is one of the most significant ways for crops to compensate for the yield loss under deadly episodes of drought stress. To prepare for the impending food crisis, the goal of developing high-yielding cultivars has been undertaken. Several technologies are being used to improve the drought tolerance mechanism to maintain the high yielding status of soybean genotypes. There is still a need to do a lot of work to develop drought tolerance soybean genotypes. The integrated use of techniques like genes transfer, QTL pyramiding, improved conventional breeding methods, genetic engineering, transcriptome, and CRISPR/Cas9 will assist the researchers in enhancing the drought tolerance in soybean. The wild genotypes of soybean should be studied to develop novel and desired traits in soybean. The episodes of drought stress will continue to increase soon; however, using improved molecular and conventional techniques would surely improve the soybean performance against drought stress.
Acknowledgement
The authors are thankful to Prof. Dr. Ziming Wu for his financial support for this study.
Funding
The research was supported by the National Natural Science Foundation of China (31760350 and 71963020), Training Program for Academic and Technical Leaders in Major Discipline in Jiangxi Province (20204BCJL22044) and Natural Science Foundation of Jiangxi (20202BABL205020), and the Jiangxi Agriculture Research System (JXARS-18).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Authors declare that they have no significant competing financial, professional, or personal interests that might have influenced the performance or presentation of the work described in this manuscript.
References
- Combined effect of deficit irrigation and potassium fertilizer on physiological response, plant water status and yield of soybean in calcareous soil. Arc. Agro. Soil Sci.. 2017;63(6):827-840.
- [Google Scholar]
- Mapping of quantitative trait loci for canopy-wilting trait in soybean (Glycine max L. Merr) Theoret. App. Gen.. 2012;125(5):837-846.
- [Google Scholar]
- Comprehensive RNA-seq analysis revealed molecular pathways and genes associated with drought tolerance in wild soybean (Glycine soja Sieb. and Zucc.) Physiol. Plant.. 2021;172:707-732.
- [Google Scholar]
- Genome-wide characterization and functional analysis of class III peroxidase gene family in soybean reveal regulatory roles of GsPOD40 in drought tolerance. Genomics. 2022;114(1):45-60.
- [Google Scholar]
- Morphological and yield response of soybean (Glycine max) to foliar application of antioxidants under drought condition. Plant Cell Biotech Mol. Biol.. 2021;22:103-110.
- [Google Scholar]
- Implications of ethylene biosynthesis and signaling in soybean drought stress tolerance. BMC Plant Biol.. 2015;15:1-20.
- [Google Scholar]
- Overexpression of PIF4 affects plant morphology and accelerates reproductive phase transitions in soybean. Food Ener. Sec.. 2021;10:e291.
- [Google Scholar]
- CRISPR/Cas9-mediated targeted mutagenesis of GmSPL9 genes alters plant architecture in soybean. BMC Plant Biol.. 2019;19(1)
- [Google Scholar]
- Ameliorating drought stress effects on soybean physiology and yield by hydrogen peroxide. Agric. Consp.. 2020;85:211-218.
- [Google Scholar]
- Yield and Quality of Two Soybean Cultivars in Response to Drought and N Fertilization. Teki. Ziraat Fakül Derg.. 2020;17:203-210.
- [Google Scholar]
- Physiology of soybean as affected by PEG-induced drought stress. Current Plant Biol/. 2020;22
- [Google Scholar]
- Interactive effect of drought and cadmium stress on soybean root morphology and gene expression. Ecotoxicol. Environ. Saf.. 2019;175:90-101.
- [Google Scholar]
- Identification of quantitative trait loci associated with canopy temperature in soybean. Scient. Rep.. 2020;10:1-11.
- [Google Scholar]
- Seed germination behavior, growth, physiology and antioxidant metabolism of four contrasting cultivars under combined drought and salinity in soybean. Antioxidants. 2022;11:498.
- [Google Scholar]
- Suppression of ERECTA Signaling Impacts Agronomic Performance of Soybean (Glycine max (L) Merril) in the Greenhouse. Front. Plant Sci.. 2021;12:644.
- [Google Scholar]
- Sensitivity of Seed Germination and seedling radicle growth to drought stress in Sesame Sesamum indicum L. Res. J. Environ. Sci.. 2011;5:557.
- [Google Scholar]
- Businesswire, 2021. Bioceres Crop Solutions Receives Approval of HB4® Drought Tolerant Soybean in Canada. Businesswire [Online]. Available at: https://www.businesswire.com/news/home/20210601005303/en/Bioceres-Crop-Solutions-Receives-Approval-of-HB4%C2%AE-Drought-Tolerant-Soybean-in-Canada.
- Identification and confirmation of loci associated with canopy wilting in soybean using genome-wide association mapping. Front. Plant Sci.. 2021;12:698116
- [CrossRef] [Google Scholar]
- Strategies to mitigate the adverse effect of drought stress on crop plants—influences of soil bacteria: A review. Pedosphere. 2021;31(3):496-509.
- [Google Scholar]
- Polygenic inheritance of canopy wilting in soybean (Glycine max (L.) Merr.) Theoret. App. Genet.. 2009;119(4):587-594.
- [Google Scholar]
- Overexpression of GmMYB14 improves high-density yield and drought tolerance of soybean through regulating plant architecture mediated by the brassinosteroid pathway. Plant Biotech. J.. 2021;19(4):702-716.
- [Google Scholar]
- Overexpression of transcription factor GmTGA15 enhances drought tolerance in transgenic soybean hairy roots and arabidopsis plants. Agronomy. 2021;11:170.
- [Google Scholar]
- Effect of drought stress on water relation traits of four soybean genotypes. SAARC J. Agric.. 2017;15:163-175.
- [Google Scholar]
- North American drought: Reconstructions, causes, and consequences. Earth-Sci. Rev.. 2007;81(1-2):93-134.
- [Google Scholar]
- Quantitative lasting effects of drought stress at a growth stage on soybean evapotranspiration and aboveground biomass. Water. 2021;13:18.
- [Google Scholar]
- DArT-based evaluation of soybean germplasm from Polish Gene Bank. BMC Res. Notes. 2021;14:1-6.
- [Google Scholar]
- Strategies to deal with drought-stress in biological nitrogen fixation in soybean. App. Soil Ecol.. 2022;172
- [Google Scholar]
- Evaluation of elite southern maturity soybean breeding lines for drought-tolerant traits. Agro. J.. 2014;106(6):1947-1954.
- [Google Scholar]
- Quantitative trait locus mapping for drought tolerance in soybean recombinant inbred line population. Plants. 2021;10:1816.
- [Google Scholar]
- Reactive oxygen species as universal constraints in life-history evolution. Proceedings of the Royal Society B: Biol. Sci.. 2009;276(1663):1737-1745.
- [Google Scholar]
- Effect of drought stress at reproductive stages on growth and nitrogen metabolism in soybean. Agronomy. 2020;10:302.
- [Google Scholar]
- Water stress, photosynthesis, seed sucrose levels and seed growth in soybean. The J. Agricu Sci.. 2004;142(1):1-8.
- [Google Scholar]
- Drought stress in grain legumes during reproduction and grain filling. J Agro. Crop Sci.. 2017;203(2):81-102.
- [Google Scholar]
- Using large-scale climate drivers to forecast meteorological drought condition in growing season across the Australian wheatbelt. Sci. The Total Environ.. 2020;724
- [Google Scholar]
- SMXLs regulate seed germination under salinity and drought stress in soybean. Plant Growth Reg.. 2022;96(3):397-408.
- [Google Scholar]
- Constitutive expression of Arabidopsis bZIP transcription factor AREB1 activates cross-signaling responses in soybean under drought and flooding stresses. J. Plant Physiol.. 2021;257
- [Google Scholar]
- Identification, inheritance and QTL mapping of root traits related to tolerance to rhizo-spheric stresses in soybean (G. max (L.) Merr.) Front. Agric. China. 2007;1(2):119-128.
- [Google Scholar]
- Medicago truncatula and Glycine max: different drought tolerance and similar local response of the root nodule proteome. J. Prot. Res.. 2015;14(12):5240-5251.
- [Google Scholar]
- Identification of agronomical and morphological traits contributing to drought stress tolerance in soybean. Aust. J. Crop Sci.. 2019;13(01):35-44.
- [Google Scholar]
- SoyBase, the USDA-ARS soybean genetics and genomics database. Nucl. Acids Res.. 2010;38:D843-D846.
- [Google Scholar]
- Expression analysis in response to drought stress in soybean: Shedding light on the regulation of metabolic pathway genes. Gen. Mol. Biol.. 2012;35(1 suppl 1):222-232.
- [Google Scholar]
- Decreasing nitrogen assimilation under drought stress by suppressing DST-mediated activation of Nitrate Reductase 1.2 in rice. Mol. Plant. 2022;15(1):167-178.
- [Google Scholar]
- Phosphorus fertilizer placement and tillage affect soybean root growth and drought tolerance. Agro. J.. 2017;109(6):2936-2944.
- [Google Scholar]
- Role of jasmonic acid and abscisic acid treatments in alleviating the adverse effects of drought stress and regulating trypsin inhibitor production in soybean plant. Aust. J. Basic. Appl. Sci.. 2009;3:904-919.
- [Google Scholar]
- Confirmation of delayed canopy wilting QTLs from multiple soybean mapping populations. Theoret. App. Genet.. 2015;128(10):2047-2065.
- [Google Scholar]
- Effects of rhizobia and arbuscular mycorrhizal fungi on yield, size distribution and fatty acid of soybean seeds grown under drought stress. Microbiol. Res.. 2021;242
- [Google Scholar]
- Exogenous melatonin induces drought stress tolerance by promoting plant growth and antioxidant defence system of soybean plants. AoB Plants. 2021;13(4)
- [Google Scholar]
- Drought tolerance of soybean (Glycine max L. Merr.) by improved photosynthetic characteristics and an efficient antioxidant enzyme activities under a split-root system. Front. Physiol.. 2019;10
- [Google Scholar]
- Photosynthetic Physiology Performance and Expression of Transcription Factors in Soybean of Water Use Efficiency Difference. Russ. J. Plant Physiol.. 2022;69:1-10.
- [Google Scholar]
- Interaction between phosphorus nutrition and drought on grain yield, and assimilation of phosphorus and nitrogen in two soybean cultivars differing in protein concentration in grains. Journal of Plant Nutrition. 2006;29(8):1433-1449.
- [Google Scholar]
- Crosstalk between phytohormones and secondary metabolites in the drought stress tolerance of crop plants: a review. Physiol. Plant.. 2021;172(2):1106-1132.
- [Google Scholar]
- Natural variation and genetic constraints on drought tolerance. Current Opinion in Plant Biol.. 2013;16(3):274-281.
- [Google Scholar]
- Yield associated traits correlate with cytokinin profiles in developing pods and seeds of field-grown soybean cultivars. Field Crops Res.. 2017;214:175-184.
- [Google Scholar]
- Rapid and surprising dieback of Utah juniper in the southwestern USA due to acute drought stress. Forest Ecol. Manag.. 2021;480
- [Google Scholar]
- Analysis of QTL–allele system conferring drought tolerance at seedling stage in a nested association mapping population of soybean (Glycine max L. Merr.) using a novel GWAS procedure. Planta. 2018;248(4):947-962.
- [Google Scholar]
- Using the RTM-GWAS procedure to detect the drought tolerance QTL-allele system at the seedling stage under sand culture in a half-sib population of soybean (Glycine max (L.) Merr.) Canad. J. Plant Sci.. 2019;99(6):801-814.
- [Google Scholar]
- Case study: cost-effective image analysis method to study drought stress of soybean in early vegetative stage. J. Crop Sci. Biotech.. 2022;25(1):33-37.
- [Google Scholar]
- In search for drought-tolerant soybean: is the slow-wilting phenotype more than just a curiosity? J. Exp. Bot.. 2020;71(2):457-460.
- [Google Scholar]
- Inter-tissue and inter-organ signaling in drought stress response and phenotyping of drought tolerance. The Plant J.. 2022;109:342-358.
- [CrossRef] [Google Scholar]
- Differential gene expression in soybean leaf tissues at late developmental stages under drought stress revealed by genome-wide transcriptome analysis. PloS One. 2012;7(11)
- [Google Scholar]
- Identification of soybean yield QTL in irrigated and rain-fed environments. Agronomy. 2021;11(11):2207.
- [Google Scholar]
- Nitrogen supply enhances the physiological resistance of Chinese fir plantlets under polyethylene glycol (PEG)-induced drought stress. Sci Rep.. 2020;10:1-8.
- [Google Scholar]
- Overexpression of Arachis hypogaea AREB1 gene enhances drought tolerance by modulating ROS scavenging and maintaining endogenous ABA content. Inte J. Mol. Sci.. 2013;14:12827-12842.
- [Google Scholar]
- Dynamic risk assessment of drought disaster for maize based on integrating multi-sources data in the region of the northwest of Liaoning Province. China. Nat. Haza.. 2013;65(3):1393-1409.
- [Google Scholar]
- Genome-wide association study of soybean seed germination under drought stress. Mol. Genet. Gen.. 2020;295(3):661-673.
- [Google Scholar]
- Engineering multiple abiotic stress tolerance in canola. Brassica napus. Fronti. Plant Sci.. 2020;11
- [CrossRef] [Google Scholar]
- Physiological response of Kalanchoe tubiflora leaves to drought stress and rewatering. J. Trop. Subtrop. Bot.. 2014;22:391-398.
- [Google Scholar]
- The development and improvement of drip irrigation under plastic film on cotton. Agric. Res. Arid Areas. 2004;22:202-208.
- [Google Scholar]
- Cold, salinity and drought stresses: an overview. Arc. Biochem. Biophy.. 2005;444(2):139-158.
- [Google Scholar]
- Physiological and molecular approaches to improve drought resistance in soybean. Plant Cell Physiol.. 2009;50(7):1260-1276.
- [Google Scholar]
- The sunflower HD-Zip transcription factor HAHB4 is up-regulated in darkness, reducing the transcription of photosynthesis-related genes. J. Exp. Bot.. 2008;59:3143-3155.
- [Google Scholar]
- Genetic Transformation to Confer Drought Stress Tolerance in Soybean (Glycine max L.) Sustainable Agriculture Reviews. 2020;45
- [Google Scholar]
- Overexpression of full-length and partial DREB2A enhances soybean drought tolerance. Agro. Sci. Biotech.. 2022;8:1-21.
- [Google Scholar]
- Genetic parameters of agronomic traits in soybean (Glycine max L. In: Merrill) genotypes tolerant to drought, AIP Conference Proceedings. AIP Publishing LLC. 2022. p. :020003.
- [Google Scholar]
- Physiological approach to decipher the drought tolerance of a soybean genotype from Brazilian savana. Plant Physiol.. 2020;151:132-143.
- [Google Scholar]
- Improvement of drought tolerance of soybean plants by using methyl jasmonate. Physiol. Mol. Biol. Plants. 2017;23:545-556.
- [Google Scholar]
- Drought tolerance responses in vegetable-type soybean involve a network of biochemical mechanisms at flowering and pod-filling stages. Plants. 2021;10:1502.
- [Google Scholar]
- Drought stress during soybean seed filling affects storage compounds through regulation of lipid and protein metabolism. Acta Physiol. Plant.. 2018;40:1-8.
- [Google Scholar]
- The transcriptional regulatory network in the drought response and its crosstalk in abiotic stress responses including drought, cold, and heat. Front. Plant Sci.. 2014;5:170.
- [Google Scholar]
- Overexpression of the GmDREB6 gene enhances proline accumulation and salt tolerance in genetically modified soybean plants. Sci. Rep.. 2019;9:1-8.
- [Google Scholar]
- Overexpression of Glycine soja WRKY20 enhances drought tolerance and improves plant yields under drought stress in transgenic soybean. Mol. Breed.. 2017;37:19.
- [Google Scholar]
- Optimization of CRISPR/Cas9 genome editing to modify abiotic stress responses in plants. Sci. Rep.. 2016;6:1-10.
- [Google Scholar]
- Recent advances in breeding for drought and salt stress tolerance in soybean. In: Jenks M.A., Hasegawa P.M., Jain S.M., eds. Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops. Dordrecht: Springer Netherlands; 2007. p. :739-773.
- [Google Scholar]
- Cytokinin-mediated source/sink modifications improve drought tolerance and increase grain yield in rice under water-stress. Plant Biotech. J.. 2011;9:747-758.
- [Google Scholar]
- Knockdown of GmSOG1 Compromises drought tolerance in transgenic soybean lines. Amer. J. Plant Sci.. 2021;12:18.
- [Google Scholar]
- Agronomic characters of early maturity soybean mutant lines. AIP Conference Proceedings. AIP Publishing LLC 2021020068
- [Google Scholar]
- Proteomic analysis of elite soybean Jidou17 and its parents using iTRAQ-based quantitative approaches. Prot. Sci.. 2013;11:1-11.
- [Google Scholar]
- Acetic acid improves drought acclimation in soybean: an integrative response of photosynthesis, osmoregulation, mineral uptake and antioxidant defense. Physiol. Plant.. 2021;172(2):334-350.
- [Google Scholar]
- Leaf Proteome response to drought stress and antioxidant potential in tomato (Solanum lycopersicum L.) Atmosphere. 2021;12(8):1021.
- [Google Scholar]
- Iron toxicity, tolerance and quantitative trait loci mapping in rice; a review. App. Ecol. Environ. Res.. 2020;18(6):7483-7498.
- [Google Scholar]
- A critical review on the improvement of drought stress tolerance in rice (Oryza sativa L.) Not. Bot. Horti Agro. Cluj-Nap.. 2020;48:1756-1788.
- [Google Scholar]
- Heavy metals stress and plants defense responses, Sustainable Soil and Land Management and Climate Change. CRC Press 2021:57-82.
- [Google Scholar]
- Somatic embryogenesis and plant regeneration from commercial soybean cultivars. Plants. 2020;9:38.
- [Google Scholar]
- QTL mapping of drought tolerance traits in soybean with SLAF sequencing. The Crop J.. 2020;8(6):977-989.
- [Google Scholar]
- Successful field performance in warm and dry environments of soybean expressing the sunflower transcription factor HB4. J. Exp. Bot.. 2020;71(10):3142-3156.
- [Google Scholar]
- Improved drought stress tolerance in Arabidopsis by CRISPR/dCas9 fusion with a Histone AcetylTransferase. Sci. Rep.. 2019;9(1)
- [Google Scholar]
- Effect of drought stress and aerosols on yield and some physiological traits of soybean (Glysin max L.) J. Plant Proc. Funct.. 2021;10:263.278.
- [Google Scholar]
- Evaluating the Effect of Cytokinin Foliar Application on Morphological Traits and Yield of Quinoa (Chenopodium quinoa Willd.) under Optimal Irrigation and Drought Stress Conditions. J. Crop Ecophysiol.. 2021;14:479-498.
- [Google Scholar]
- Breeding and molecular approaches for evolving drought-tolerant soybeans. Plant Stress Biology. Springer 2020:83-130.
- [Google Scholar]
- Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants. 2021;10:259.
- [Google Scholar]
- Expression of the Arabidopsis AtMYB44 gene confers drought/salt-stress tolerance in transgenic soybean. Mol. Breed.. 2012;29(3):601-608.
- [Google Scholar]
- RNA-Seq Atlas of Glycine max: a guide to the soybean transcriptome. BMC Plant Biol.. 2010;10(1):160.
- [Google Scholar]
- Over-expression of Arabidopsis ORANGE gene enhances drought stress tolerance through ABA-dependent pathway in Arabidopsis thaliana. Plant Growth Reg.. 2022;96(1):91-101.
- [Google Scholar]
- Proteomics unravel the regulating role of salicylic acid in soybean under yield limiting drought stress. Plant Physiol. Biochem.. 2018;130:529-541.
- [Google Scholar]
- Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot.. 2002;53:1305-1319.
- [Google Scholar]
- Transcriptional regulation of drought response: a tortuous network of transcriptional factors. Front. Plant Sci.. 2015;6:895.
- [Google Scholar]
- Inheritance and mapping of drought tolerance in soybean at seedling stage using bulked segregant analysis. Plant Gen. Res.. 2020;18(2):63-70.
- [Google Scholar]
- Foliar application of iron, zinc, and manganese nano-chelates improves physiological indicators and soybean yield under water deficit stress. J. Plant Nutrit.. 2020;43(18):2740-2756.
- [Google Scholar]
- Genetic diversity and genomic strategies for improving drought and waterlogging tolerance in soybeans. J. Exp. Bot.. 2017;68:1835-1849.
- [Google Scholar]
- The numbers game of soybean breeding in the United States. Crop Breed. App. Biotech.. 2021;21
- [CrossRef] [Google Scholar]
- Gm WRKY 27 interacts with Gm MYB 174 to reduce expression of GmNAC29 for stress tolerance in soybean plants. The Plant J.. 2015;83(2):224-236.
- [Google Scholar]
- Two homologous LHY pairs negatively control soybean drought tolerance by repressing the abscisic acid responses. New Phytol.. 2021;229(5):2660-2675.
- [Google Scholar]
- Indigenous plant species and invasive alien species tend to diverge functionally under heavy metal pollution and drought stress. Ecotoxicol. Environ. Safety. 2020;205
- [Google Scholar]
- Comprehensive identification of drought tolerance QTL-allele and candidate gene systems in Chinese cultivated soybean population. Int. J. Mol Sci.. 2020;21:4830.
- [Google Scholar]
- Effects of PGPR bioinoculants, hydrogel and biochar on growth and physiology of soybean under drought stress. Comm. Soil Sci. Plant Anal.. 2022;53:826-847.
- [Google Scholar]
- GmWRKY54 improves drought tolerance through activating genes in abscisic acid and Ca2+ signaling pathways in transgenic soybean. The Plant J.. 2019;100(2):384-398.
- [Google Scholar]
- Modelling transpiration, soil evaporation and yield prediction of soybean in North China Plain. Agric. Water Manag.. 2015;147:43-53.
- [Google Scholar]
- Drought stress has transgenerational effects on soybean seed germination and seedling vigor. PLoS One. 2019;14(9)
- [Google Scholar]
- The urban heat island of the north-central Texas region and its relation to the 2011 severe Texas drought. J. App. Meteorol. Climatol.. 2013;52:2418-2433.
- [Google Scholar]
- Risk assessment of hypertension in steel workers based on LVQ and Fisher-SVM deep excavation. Ieee Access. 2019;7:23109-23119.
- [Google Scholar]
- Physiological response of flax seedlings with different drought-resistances to drought stress. Acta Agric. Boreali-Sinica. 2019;34:145-153.
- [Google Scholar]
- Recent changes in the drought of China from 1960 to 2014. Int. J. Climatol.. 2020;40(7):3281-3296.
- [Google Scholar]
- Root system architecture, physiological and transcriptional traits of soybean (Glycine max L.) in response to water deficit: A review. Physiol. Plant.. 2021;172:405-418.
- [Google Scholar]
- GmNFYA13 improves salt and drought tolerance in transgenic soybean plants. Front. Plant Sci.. 2020;11:1584.
- [Google Scholar]
- Proteomic analysis of drought stress response mechanism in soybean (Glycine max L.) leaves. Food Sci. Nutrit.. 2021;9(4):2010-2020.
- [Google Scholar]
- GmNAC8 acts as a positive regulator in soybean drought stress. Plant Sci.. 2020;293
- [Google Scholar]
- QTLs and epistasis for drought-tolerant physiological index in soybean (Glycine max L.) across different environments. Caryologia. 2014;67(1):72-78.
- [Google Scholar]
- The importance of slow canopy wilting in drought tolerance in soybean. J. Exp. Bot.. 2020;71(2):642-652.
- [Google Scholar]
- Molecular markers improve abiotic stress tolerance in crops: a review. Plants. 2020;9:1374.
- [Google Scholar]
- A comparative proteomic study of drought-tolerant and drought-sensitive soybean seedlings under drought stress. Crop Past. Sci.. 2016;67:528-540.
- [Google Scholar]
- GmLCLs negatively regulate ABA perception and signalling genes in soybean leaf dehydration response. Plant, Cell Environ.. 2021;44:412-424.
- [Google Scholar]
- A soybean EF-Tu family protein GmEF8, an interactor of GmCBL1, enhances drought and heat tolerance in transgenic Arabidopsis and soybean. Int. J. Biol. Macromol.. 2022;205:462-472.
- [Google Scholar]
- Dissection of genetic overlap of drought and low-temperature tolerance QTLs at the germination stage using backcross introgression lines in soybean. Mol. Biol. Rep.. 2012;39:6087-6094.
- [Google Scholar]
- Identification of genes for drought resistance and prediction of gene candidates in soybean seedlings based on linkage and association mapping. The Crop J 2021
- [CrossRef] [Google Scholar]
- The GmXTH1 gene improves drought stress resistance of soybean seedlings. Mol. Breed.. 2022;42:1-18.
- [Google Scholar]
- The origin and evolution of cultivated soybean (Glycine max (L.) Merr.) Scient. Agric. Sinica. 2004;37:954-962.
- [Google Scholar]
- Advances and prospects: biotechnologically improving crop water use efficiency. Critical Rev. Biotechnol.. 2011;31:281-293.
- [Google Scholar]
- Yield estimation of soybean breeding lines under drought stress using unmanned aerial vehicle-based imagery and convolutional neural network. Biosy. Engin.. 2021;204:90-103.
- [Google Scholar]
- Genome-wide analysis of the soybean DREB gene family: Identification, genomic organization and expression profiles in response to drought stress. Plant Breed.. 2020;139:1158-1167.
- [Google Scholar]
- Plant survival under drought stress: Implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol. Res.. 2021;242:126626
- [Google Scholar]
- The effect of micronutrient foliar application on yield, seed quality and some biochemical traits of soybean cultivars under drought stress. J. Plant Nutrit.. 2019;42:2715-2730.
- [Google Scholar]




