

Translate this page into:
Himalayan ibex (Capra ibex sibirica) habitat suitability and range resource dynamics in the Central Karakorum National Park, Pakistan
⁎Corresponding author at: State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China. Tel.: +86 13609923012. chenxi@ms.xjb.ac.cn (Chen Xi)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
The study investigates Himalayan ibex (Capra ibex sibirica) and their range resource condition within the preferred habitat in the Central Karakoram National Park, Pakistan. We apply ecological niche factor analysis (ENFA) using 110 ibex sighting data and 6 key biophysical variables describing the habitat conditions and produce habitat suitability and maps with GIS and statistical tool (BioMapper). The modeling results of specialization factor shows some limitation for ibex over the use of slope, elevation, vegetation types and ruggedness. The habitat area selection for the ibex is adjusted to the ibex friendly habitat available conditions. The model results predicted suitable habitat for ibex in certain places, where field observation was never recorded. The range resource dynamics depict a large area that comes under the alpine meadows has the highest seasonal productivity, assessed by remote sensing based fortnightly vegetation condition data of the last 11 years. These meadows are showing browning trend over the years, attributable to grazing practices or climate conditions. At lower elevation, there are limited areas with suitable dry steppes, which may cause stress on ibex, especially during winter.
Keywords
Himalayan ibex
Species modeling
Habitat assessment
Range resources
Ecological niche factor analysis (ENFA)

1 Introduction
Himalayan ibex (Capra ibex sibirica) range is spread across the mountains of Pakistan, China, India, Afghanistan, Kyrgyzstan, Kazakhstan, Uzbekistan, Mongolia, Russia and Tajikistan (Shackleton, 1997). This species is listed as the least concerned and gets limited attention hence its unaware population trends throughout its distribution range cannot rightly define the status (IUCN, 2012). In the Hindukush–Karakoram–Himalayan (HKH) region it inhabits the relatively dry mountains between 2000 m and 5000 m elevations (Muhammad Zafar et al., 2014). Within Pakistan, it is present predominantly in the Hunza and Skardu districts of Gilgit-Baltistan province, and Dir, Swat, Kohistan and Chitral districts of Khyber Pakhtunkhwa province (Roberts, 1977; Schaller, 1977). Mir of Hunza estimated 1000 ibex population for Hunza (Roberts, 1977), whereas Rasool (1990) contradicted that ibex population of Gilgit-Baltistan declined to below 600 primarily because of illegal hunting and poaching.
Himalayan ibex mainly occupies rocky mountainous regions, both open meadows and cliffs, tracking out to low elevations during winter (Mirza, 1998; Fedosenko and Blank, 2001). The species does not enter densely forested areas, in summer or hot days crave for shaded areas under rocks or plants, live and remain near steep and escape terrain (Fedosenko and Blank, 2001). The diet of ibex consists of alpine plump and sages. It is crepuscular in feeding, foraging in evenings and mostly in early morning hours (Fedosenko and Blank, 2001). Ibex lives in small groups (6–30 animals) varying considerably in size, rarely in herds of >100 animals (Reading et al.,1995, 1997, 1999; Fedosenko and Blank, 2001).
The presence of species depends upon the specific environmental conditions that enable it to survive and reproduce (Marzluff and Ewing, 2001). Understanding on factors influencing its existence is basic requirement for the assessment of the species distribution and devising efficient species conservation strategies (Wein, 2002). Spatial (habitat) and temporal (seasons) variables define the ecological niche of a certain species within a given environment.
Multivariate statistical techniques, like Discriminant analysis, are the most popular tools for representing niche geometry and understanding the habitat–species relationship in the natural world (Dueser and Shugart, 1978; Marnell, 1998). These techniques have been widely used in defining the specific niches of species within a given environment (Mwangi and Western, 1997) based on topographical, habitat and other variables.
Himalayan rangelands support a large livestock population along with providing habitat to diverse wild ungulate species. Bagchi et al. (2004) studied competition of Himalayan ibex with livestock in the Trans-Himalayan Mountains to suggest high levels of resource use. Similar competition can exist between different wild species which can possibly out-compete ibex. For HKH region scientific data on range resources are limited which hinders the sustainable management of these resources both for livelihood improvement and nature conservation (Singh et al., 2009). The need for up-to-date information has long been recognized and stimulated the use of earth data using remote sensing techniques, which has become a universal and familiar instrument for natural resource management and implementation (Philipson and Lindell, 2003; Stow et al., 2004). Combined information from low-altitude satellite sensors and remote sensing offer an optimal path for understanding pattern and process related to rangeland condition in the area. The multi-temporal and multi-spectral data acquired by various satellite sensors are used to identify, map and monitor rangelands vegetation. Normalized difference vegetation index (NDVI) is an important method for the earth surface vegetation classification. Its chronological deviation is of great significance for revealing the advancement of earth system on a regional scale (Li and Shi, 2000). Calculation of temporal series of NDVI is very important to the remote sensing (RS) of vegetation phenology and to extract numerical records linked to the dynamics of vegetation (Hall-Beyer, 2003; Pettorelli et al., 2005).
A few ecological niche factor analysis (ENFA) based scientific studies have been conducted on large mammals in the western countries (Hirzel and Perrin, 1998; Hirzel et al., 2001; Dettki et al., 2003; Traill and Bigalke, 2006; Praca and Gannier, 2008; Praca et al., 2009) and in Karakoram Pamir mountain of China and Pakistan (Khan et al., 2012) but the species (Siberian ibex) is yet to fully investigate the HKH mountain region of Pakistan. Awan et al. (2004) studied grazing competition between wild (urial; Ovis viginei) and domestic ungulates (goats and sheep) in Salt range (Himalayan foot-hills) of Pakistan. The study focused on diet composition and competition, and did not consider other biophysical variables (aspect, slope, elevation, distance to escape terrain, ruggedness, landcover, etc.) required for description of urial habitat. Arshad et al. (2012) predicted potential habitat of the Kashmir markhor (Capra falconeri cashmiriensis) in order to understand the underlying environmental conditions by map using ENFA in the Chitral Gol National Park, Pakistan.
Awan et al. (2004) and Arshad et al. (2012) implemented ENFA for their specific study within Pakistan for food competition with domestic and wild life species. Here we implement ENFA for habitat suitability modeling and habitat range dynamics. ENFA produces habitat suitability maps that indirectly demonstrate species potential distribution within the study area on presence only dataset. This compares the species mean distribution (response of the ecological variables) with the global mean (ecological characteristics of the whole study area) distribution. It is difficult for the related species to co-exist and share the same ecological niche, which mostly results in the competitive exclusion of one of the co-existing species (Praca and Gannier, 2008). Habitat modeling increases understanding regarding species distribution and its ecological interaction with environmental variables. This has been very effective to determine species ecological niches as well as for defining and managing protected areas (Praca et al., 2009).
The study was therefore specially designed as an attempt to investigate the current state of rangeland resource within the preferred habitat of Siberian ibex (Capra ibex sibirica) in the Central Karakoram National Park (Pakistan), using only the species presence data, for habitat modeling in the ENFA model to produce habitat suitability maps and eco-geographical variables characterizing key habitat conditions, for future improved management of Siberian ibex in Central Karakoram National Park, Pakistan.
2 Material and methods
2.1 Study area
Central Karakoram National Park (CKNP; notified in 1993) is the largest national park of Pakistan, with a core area of approximately 10,000 km2. It has 17 main valleys holding a human population of ∼212,000 and a total area covering all the valleys (including buffer communities) is 19,000 km2 (Fig. 1). It is a highly glaciated part of the Karakoram with altitudes ranging between 1500 m and 8611 m (at the summit of K-2) above mean sea level (asl). The park falls within administrative boundaries of the five districts of Gilgit-Baltistan, viz. Ghanche, Skardu, Shigar, Gilgit and Nagar. The varied range of altitude makes CKNP a unique hub of diverse ecosystems harboring several threatened species of flora and fauna, which are indigenous to the Karakoram. CKNP is home to 33 mammal species (12 carnivores, 9 rodents, 5 artiodactyls, 5 chiropterans, 2 lagomorphs) including, Himalayan ibex (Capra ibex sibirica), snow leopard (Unciauncia), Himalayan lynx (Felis lynx), musk deer (Moschus chrysogaster), black bear (Ursus thebatinus), Ladakh urial (Ovisvingei) and blue sheep (Pseudois nayaur). Pipistrelle, Asian barbestelle, desert long eared bat and tube nosed bat, Indian gray wolf, Kashmir or hill fox, Tibetan red fox, Alpine weasel, common otter and Lynx are also present within the park limits (Khan, 2013).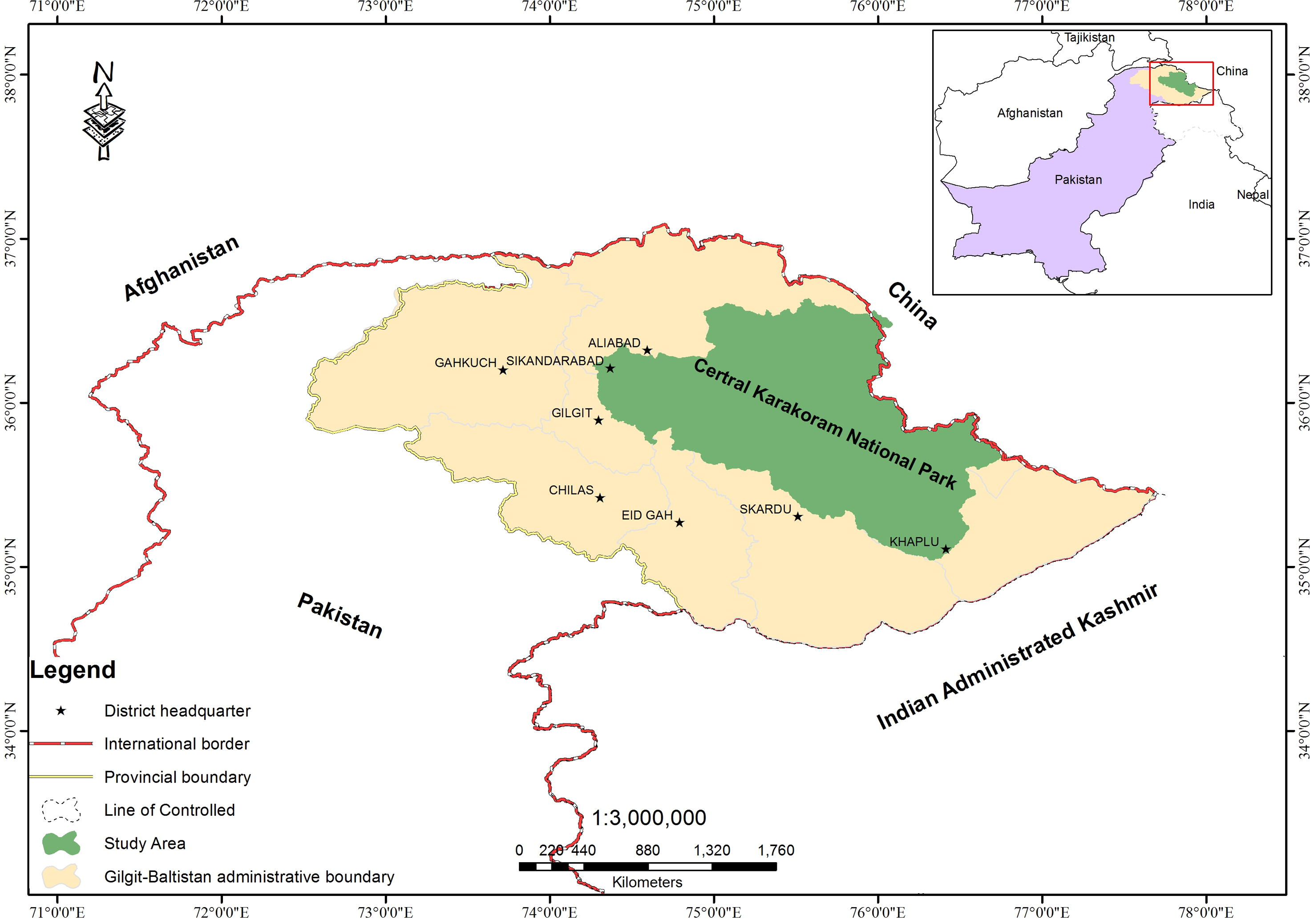
Location map of CKNP.
The study is structured in two main parts which include calculating habitat suitability maps and assessment of range resource dynamics. Later, resource availability is compared with the potential distribution of the species to assess the habitat status of the ibex in CKNP as shown in the study scheme below (Fig. 2).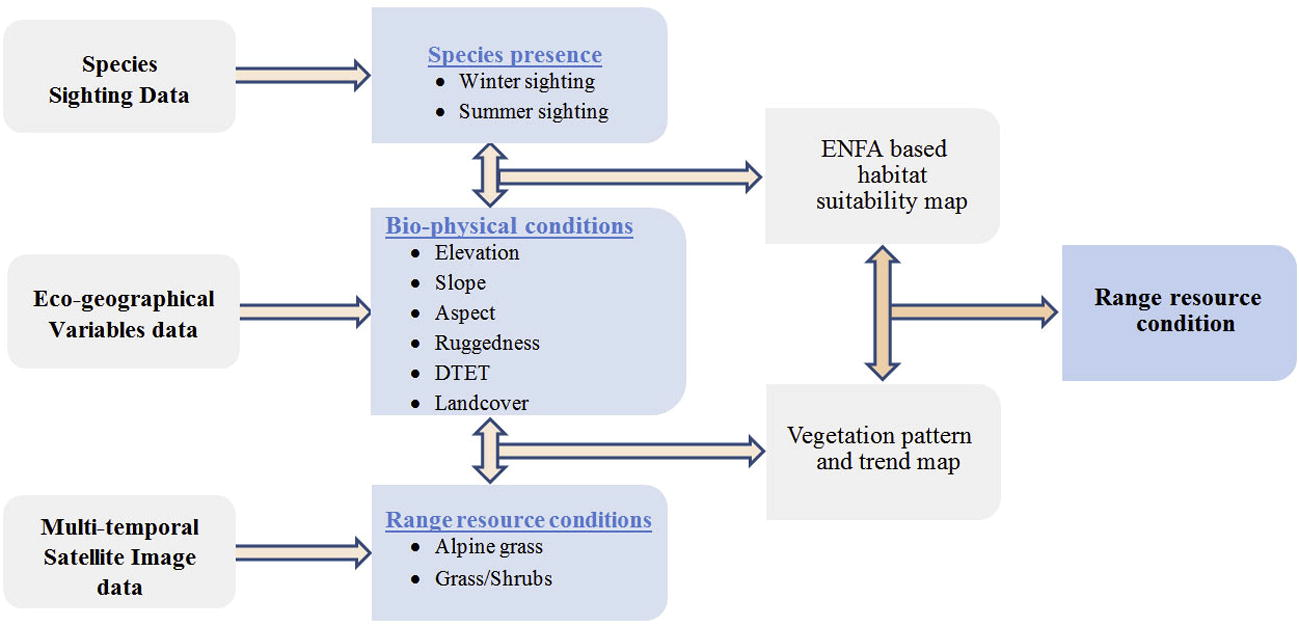
Study scheme/flow diagram.
2.2 Species sighting
Seasonal surveys (summer and winter) were carried out in most of the valleys of CKNP to collect ibex data, using field encoding forms, describing direct sighting of the species and fixed-point direct count method, mostly on randomly followed transects from high vantage fixed points along ridgelines (Aryal et al., 2010). Direct sightings (Fig. 3) as well as feces were collected as an evidence of the presence of species.
Himalayan ibex herd (left) in alpine meadows and ibex in escape terrain (right) in national park.
The ibex field data of ‘presence data, consisting of 110 point locations of herds on 30 × 30 m grid cells were collected from the study area of three years (2010–2012). A herd was taken as the basic unit of measurement for ibex field data. For each observation, geographical position, elevation, bearing angle, distance from observation point, habitat condition and topography of the ibex were recorded across seasons (summer and winter). Also some additional information was recorded; time of day, ibex herd activities, foraging and pattern of grouping. Winter data were collected to observe the maximum group of mixed ibex species during the rut season (December–January). Summer data were collected during the period (May–July) of lambing to observe the kid and female ratio and also their movement in habitat. All the records were taken during morning and evening time, when ungulates are more active. In addition daylong observations were also gathered from fixed vantage point to record their specific foraging behavior and other group activities. The ibex field data were carried out from fixed vantage points (visual searching) where the maximum observation area could be scanned by a group of observers. Double counting of animals was removed from the dataset after confirming the herd size and structure and the timings of movement from one pasture to another (Arshad et al., 2012). The ibex were recognized and recorded on the basis of their age and sex (Schaller, 1977). The herds are distinct from one another on the bases of distance, grouping size and type of species within the group. A group was defined as when there was more than one animal.
In order to record field data, A3 sized base maps of ASTER satellite data (False color composites, FCCs) with topographic data of survey of Pakistan (SoP) sheets of scale 1:50,000 scales were used. Later data were processed and converted into GIS layers (30 × 30 m) grid cells having species presence using projected coordinate system (UTM 43N) with WGS 1984 projection, for the purpose of data compatibility. Furthermore species presence data were converted into Boolean format for the purposes of operating suitability model. Species presence data and eco-geographical variable grids were calculated through this resolution and extent. Subsequently converting into Idrisi (.rst) format using IDRISI Taiga (16.03) tool (Eastman et al., 2009) to solve compatibility issue of overlaying maps, further analyses were done in Bio mapper 4.0 tool (Hirzel et al., 2007).
2.3 Eco-geographical variables
List of variables were developed and collected field information as well as extracted habitat variables from the GIS/RS techniques were categorized into two classes. (1) Topographic variables (elevation, slope, aspect, terrain ruggedness and distance to escape terrain), and (2) physio ecological variables (land cover classes and vegetation types), based upon their importance to Himalayan ibex habitat.
ASTER 30 m digital elevation model (DEM) was procured from the official website of USGS geo-portal (www.wist.echo.nasa.gov). Like elevation, slope, aspect, terrain ruggedness and distance to escape terrain calculating using ArcGIS ‘spatial analyst tool’ (ESRI, 2011). Further it was processed to extract eco-geographical variables (EGV). The aspect was categorized into eight classes: i.e. east, west, north, south, north-east, north-west, south-east and south-west directions. A slope map was derived in degrees, for the purpose of specific position modeling requirements of the species. Ruggedness was calculated in ArcGIS (spatial analyst distance tool) using ArcView script (ESRI) by overlapping both aspect and slope >45 in path distance tool. Ruggedness or vector ruggedness measure (VRM) uses three-dimensional diffusion of orthogonal vectors to planar surfaces on a landscape (Arshad et al., 2012). Distance to escape terrain (DTET) was computed using ArcSpatial Analyst and ArcHydro tools in ArcGIS. Ridges were extracted from the DEM, flow accumulation and flow direction grids were calculated during the watershed delineation process in ArcGIS. Finally, ridge features were delineated using threshold values i.e. flow amassing value = 0, represents the ridge lines. Later these ridge lines were overlapped with steep slopes (>45 degree) to produce escape terrain in path distance tool of ArcGIS. Furthermore spatial analyst tool was used to calculate the distance from one escape terrain to another and the distance from the ridgeline with slope greater than 45°. The final result of these grids shows the distance to escape terrain.
Land cover of the study area developed using Landsat TM images 30 × 30 m resolution of 2009 with supervised classification in ERDAS Imagine 10 (Geosystems, 2010) was used for (ENFA) modeling. Afterward the entire datasets on species observation and eco-geographical variables (EGVs) were converted into IDRISI file format with same resolution, extent and projection for further use as input for ENFA (BioMapper) tool.
2.4 Ecological niche factor analysis
Ecological niche factor analysis (ENFA) compares the indication of animal presence to the suite of available resources in the environment by conducting factor-analysis to quantify the relationship of biotic and abiotic variables to the species presence data. The first factor is marginality, which defines mean habitat in the study area how far the species optimum is from. The second factor is specialization which is sorted by reducing the extent of clarified alteration; which defines how the species is specialized in the study area by position to the available range of habitat. Numerically the marginality vector corresponds to the vector connecting the centroid (i.e., the mean) of the distribution of availability weights to the centroid of the distribution of utilization weights. The specialization is a measure of habitat selection on a particular direction of the ecological space. Only a few of the first factors clarify the main portion of the whole information (Hirzel et al., 2001, 2002, 2006; Hirzel and Guisan, 2002; Hirzel and Arlettaz, 2003; Praca et al., 2009). A combination of marginality and specialization factors calculated for each cell on the map and overall habitat suitability index is determined. Habitat suitability (HS) is scaled to values ranging from 0% (unsuitable) to 100% (optimal) is computed and indicated how the grouping of factors of a single prison cell favors the requirements of the species studied. By comparing MF and SF factors to observed species distribution and eco-geographical variables distribution in the study area. Habitat suitability (HS) maps were calculated (Hirzel et al., 2002) in BioMapper 4.0 tool. Later based on the threshold values four habitat suitability classes (unsuitable, marginal, suitable and optimal) were defined (Sattler et al., 2007).
2.5 Range resource assessment
To assess the range resource distribution, supervised classification of Landsat TM images was performed to identify overall land cover of the area. On the basis of elevation threshold land cover map (the range lands resources) was classified into three classes, as Alpine grasses, Sub-alpine grasses, and sub-alpine dry steppe. For the range resource trend analysis, the normalized difference vegetation index (NDVI) was calculated. The moderate resolution sensor (MODIS) satellite terra images at 16-days’ interval (MOD13Q1) with spatial resolution of 250 m were used. For NDVI 16-days of total 253 images composite, covering 11 years (2001–2011) were used in the analyses. Freely available data were exploited the geoportal of the Land Processes Distributed Active Archive Center (LPDAAC), of Earth Resources Observation and Science Center (EROS) (www.lpdaac.usgs.gov), U.S. Geological Survey (USGS). Pure grasslands patches were identified by analyzing 11 years’ vegetation dynamics. Within these pure patches, mean seasonal integrand NDVI map was produced to see the spatial distribution of the productivity. To identify the change greening or browning trends pertaining to the vegetation was assessed through seasonal nonparametric Mann–Kendal (MK) test (Neeti and Eastman, 2011).
3 Results and discussion
3.1 Habitat suitability
ENFA, based on 110 presence cell and 6 eco-geographical variables, suggests that the model fits the continuous Boyce index (0.92 ± 0.03, mean ± SD; close to hypothetical maximum, i.e., 1) with a monotonically increasing line P/E curve (Fig. 4). The results of Table 1 show that the marginality of ibex had 0.22 and specialization 1.38 with a tolerance coefficient of 0.72 (1 indicates no specialization at all). The model result shows the requirements of the species vary from one habitat to the other and depend upon the average condition of available habitat condition in the study area. The factor of specialization specified some precise habitat requirements of the ibex, but still it covered a wider niche. The marginality axis proposes that higher elevations needed by ibex (coefficient of 0.81) but in winter it prefers low elevations. It shows a close association with moderately steep slope (0.76). Ibex showed limits to different land cover classes during different seasons and avoided mixed forests, conifer forests, agriculture and human habitations areas and bare rocks. The specialization axes explained that ibex restricts its habitat to steep slopes (0.76) in order to avoid predators and also desired high elevations (0.81). The results (Table 1) also show limited ibex species presence to east to northeast aspects (ranging from 0.07 to 0.15) and strong relationship with escape terrain (0.61). High ruggedness (0.70–0.76) is also a significant factor that has influenced the presence of ibex in the area.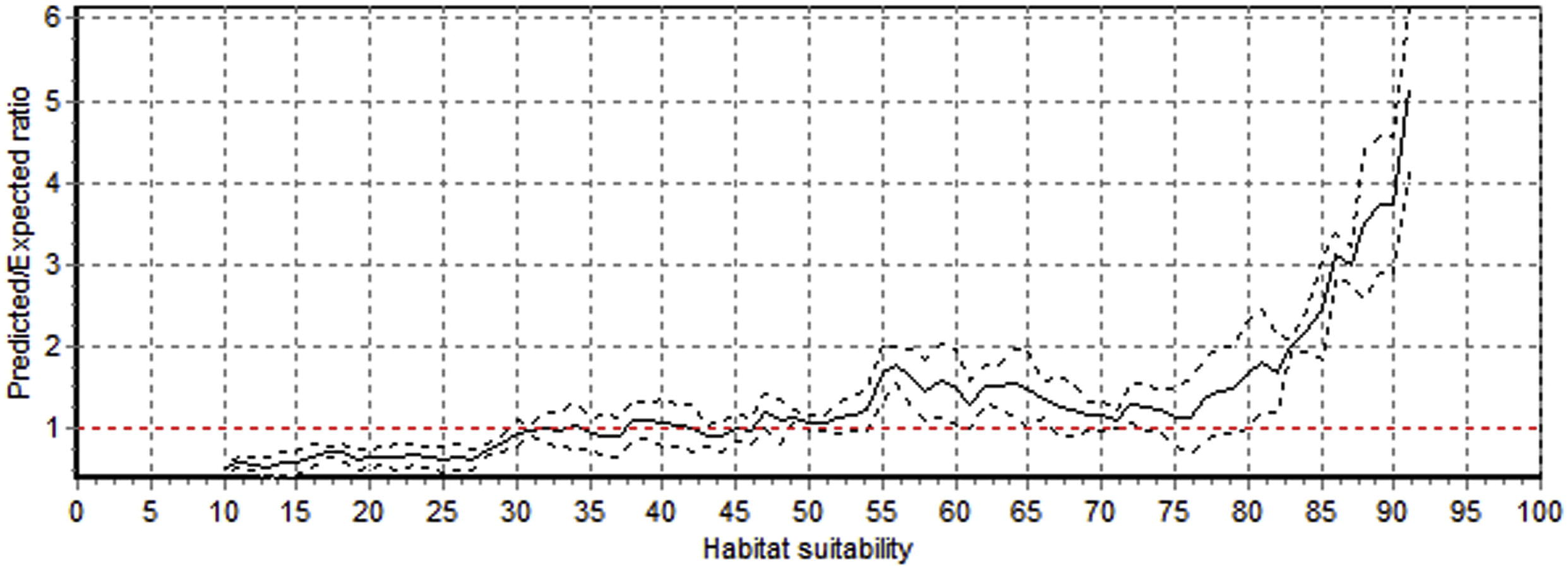
Predicted to expect (P/E) ratio curve against different habitat suitability levels for Himalayan ibex in CKNP .Solid line represents Boyce continuous curve, and dotted lines standard deviation. Red dashed line (P/E = 1) indicate habitat threshold (P/E = 1) indicate habitat threshold (P/E = 1; >1 suitable, <1 not suitable).
Eco-geographical variables
Factor 1 marginality (37)
Factor 2 specialization1 (22)
Factor 3 specialization2 (14)
Factor 4 specialization3 (11)
Aspect
−0.01
−0.06
0.15
−0.07
Elevation
0.81
0.39
−0.02
0.76
Distance to escape terrain
0.07
−0.23
−0.61
−0.14
Land cover
−0.58
−0.61
−0.55
−0.61
Terrain ruggedness
−0.03
−0.23
−0.70
0.76
Slope
0.08
0.03
0.20
−0.06
In general, the results suggest that habitat conditions are adequate to ungulate requirements. The results of specialization factor displayed some limitation for ungulates over the use of slope, elevation, vegetation types and ruggedness but the habitat selections quiet harmonized with the habitat situations available in the study area. The data were collected in two seasons (winter & summer) over a period of three years. This specified that, the species used habitat all over the year in order to precisely forecast the distribution and classify habitat suitability areas of the species. Fig. 5 clearly mention that field observation with habitat suitability model predicted suitability classes. But in some cases the species was present where the optimal and suitable classes have been predicted in the result maps. Strubbe and Matthysen (2008) observed that this may be possibly due to the mountain ungulates spread to higher suitable zones in the surroundings.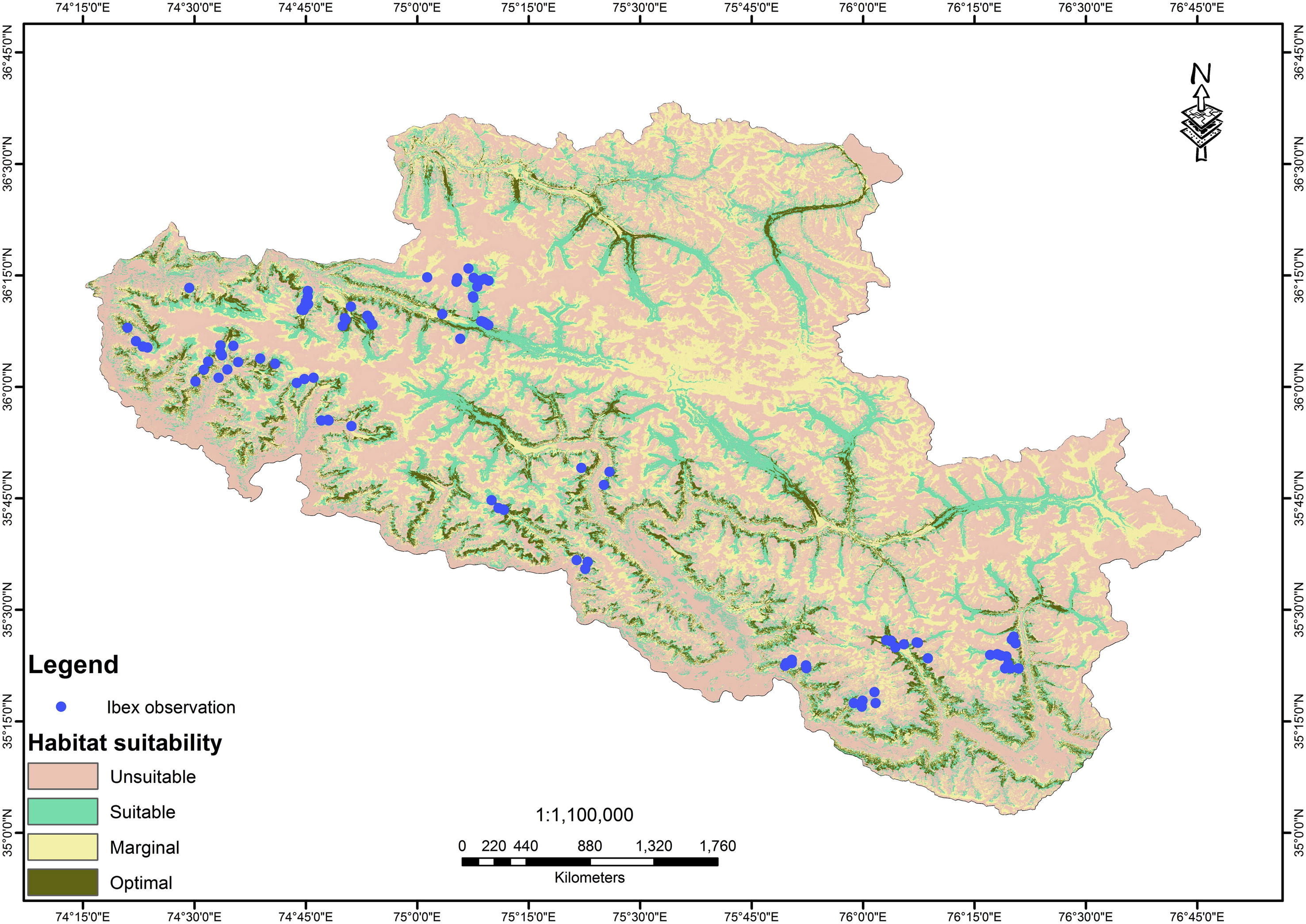
Habitat suitability map of Himalayan ibex.
The three indices are used for habitat suitability model validation which include Boyce index (B), contrast validation index (CVI) and absolute validation index (AVI). According to results of the model their respective values are (Mean ± SD) AVI: 0.483 ± 0.219, CVI: 0.209 ± 0.204, and B: 0.632 ± 0.404. These results indicate that the model evaluation is better than chance, it is purely random model and explaining a very good predictive power. These values also indicated that the model expected high habitat suitability and monotonic increase represents the very good predictive model.
3.2 Range resource dynamics
Based on the land cover assessment of Landsat images of 2010, the total vegetation cover in the CKNP is 12.5% (2,291 km2), of which 3% is dry steppe between the elevation of 2000–2600 m, 13% sub-alpine (2600–3500 m) and 62% alpine meadow (>3500 m) above sea level (asl) respectively. Around 22% of the vegetation is non-range vegetation of agriculture, forest and woody scrub <2000 m (asl; Fig. 6).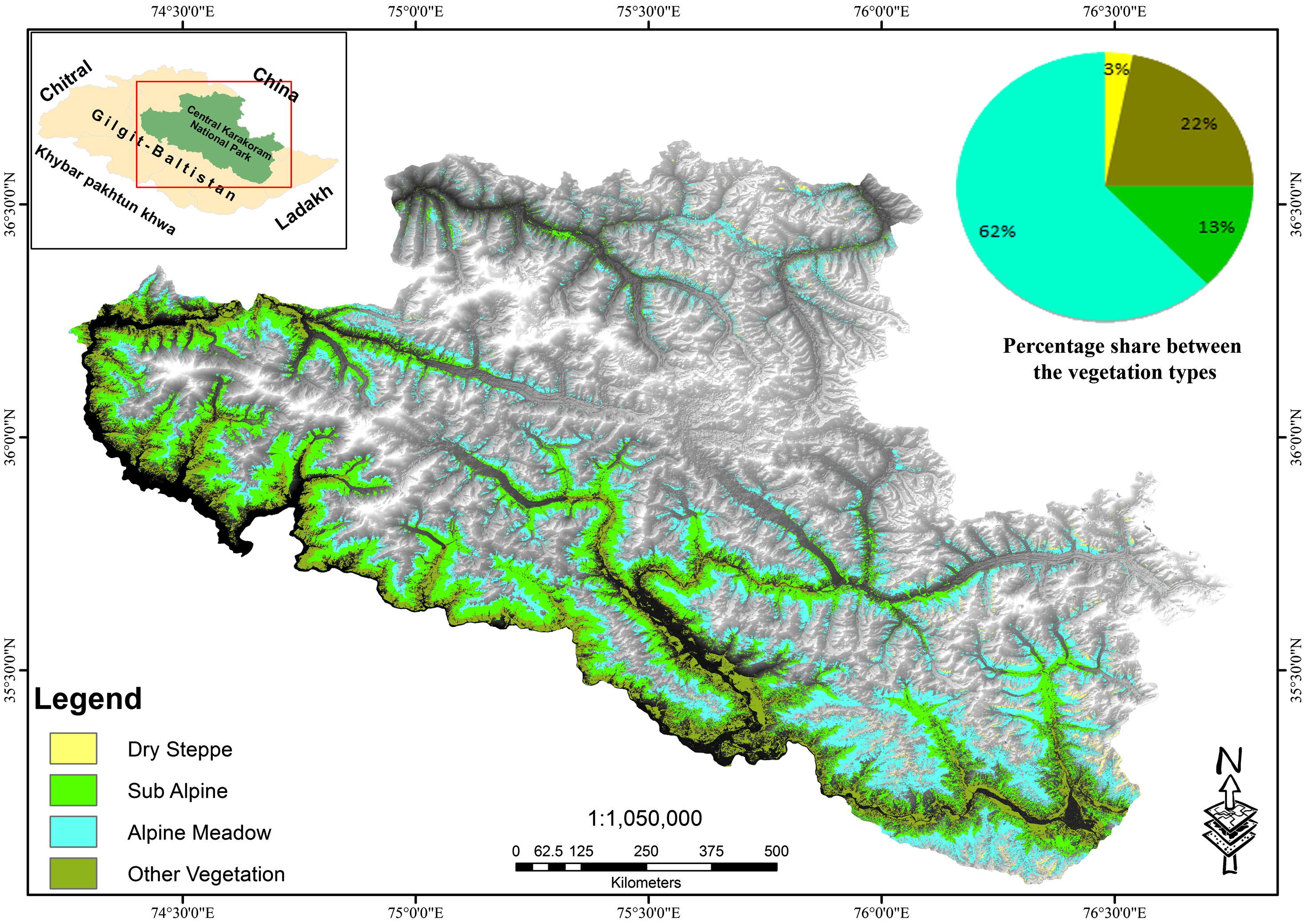
Range resource distribution in CKNP.
The dry steppe vegetation has sparsely distributed grasses and bushes of Atremesia spp. and Krascheninnikovia (Pamirian winterfat), mostly used by grazing livestock. The annual rainfall ranges between 130 and 160 mm, without a specific rainy season. The sub-alpine zone is present along the streams at upper slopes. The vegetation on this zone comprises deciduous, evergreens species and poa-grasses. The sub-alpine scrub is highly important for livestock and mountain ungulates (including ibex), being the summer grazing area. Vegetation includes birch, juniper, fir, willow and pine forests along with green alpine pastures on plains and gentle slopes with common vegetation species; common juniper, mertensia, potentilla desertorum and Elegant Willow. Xerophytic grasses appear during July and August. The landscape is characterized by large glaciers, sheer cliffs and boulders with closed sedge mats confined to shady areas.
Spatial range resource productivity is assessed in terms of season integrated NDVI of pure grasslands pixels which actually reflects photosynthetic activity and can be taken as proxy to total seasonal productivity. The productivity has positive association with elevation. Alpine meadow has higher productivity in comparison to lower elevation pastures, attributed to high precipitation and snow melt. The mean productivity values of dry steppe, sub-alpine and alpine meadows are 33,370, 45,250, and 58,300, kg/hector respectively (Fig. 7).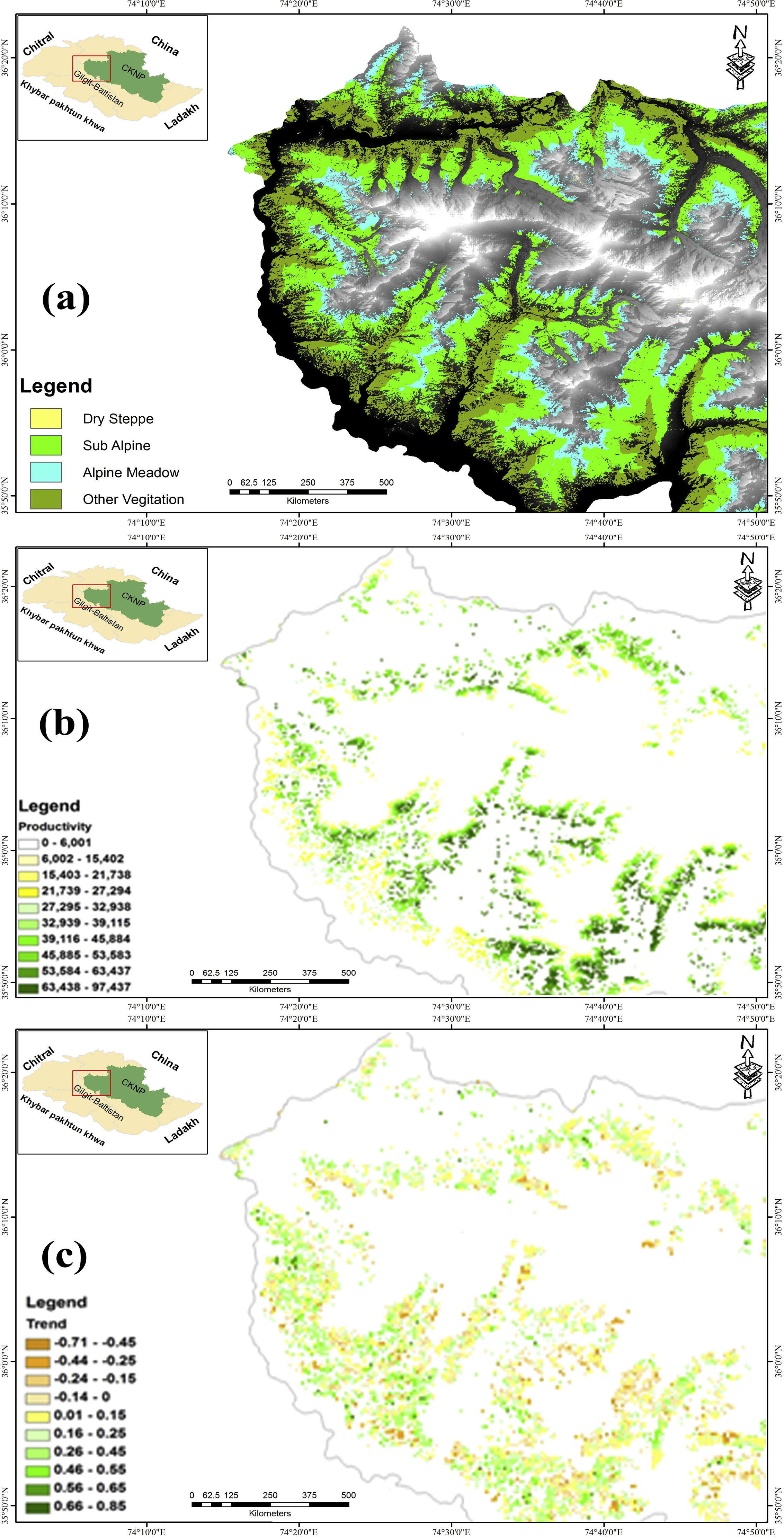
Range resource dynamics in the western part of CKNP (a) rangelands vegetation type (b) seasonal productivity (c) greening and browning trend.
Temporal change trend, based on Mann–Kendal test using 11 years fortnightly vegetation index of pure grasslands pixels within the range resources reveals that high alpine meadows have high browning trend in comparison to dry steppe pastures (Fig. 7). In the dry steppe 82% of pure pixel showed greening trend, 12% showed browning trend and 6% remained stable; in the sub-alpine region 70% of pure pixel showed greening trend, 27% showed browning trend and 6% remained stable; in the alpine meadows 60% of pure pixel showed greening trend, 39% showed browning trend and 1% remained stable. The alpine pastures are considered as fragile ecosystem within the area, where the habitat is under heavy grazing pressure. Previous studies carried out by various researchers in the region have shown declining productivity and bio-diversity in alpine and subalpine ranges (MACP/IUCN, 2001). These communal lands are being exploited for immediate use rather than long term benefit.
3.3 Species habitat preference
Rangeland resource distribution was compared with species preference for different habitats in the study area where results showed abundance of grazing grounds in alpine meadows specially for summer season followed by limited foraging resources available in the dry steppe at lower elevations for winter feeding (Table 2), which may intensify competition of the species with domestic livestock being grazed in same areas. Some of the wild herbivores may out compete in this process, while increasing the chance of disease transmission from livestock to wild animals and vice versa.
Optimal
Suitable
Marginal
Unsuitable
Dry steppe
0
1100
2000
5209
Sub alpine
2182
3800
12,194
11,631
Alpine meadow
31,215
20,860
16,396
74,574
In terms of temporal trends, rangeland resources seemed stable or improving at lower altitudes whereas, alpine pastures had a higher browning trend at higher elevations. Degradation of alpine and sub alpine pastures may directly impact health and productivity of the wild and domestic animals, and jeopardize local economy if not managed timely.
4 Conclusion
Traditionally, the mountain communities of Pakistan, and more specifically those living in buffer zone valleys of CKNP of Gilgit-Baltistan, having limited arable land (<2%, n = 72,496 km2) enjoy an agro-pastoral living pattern. Subsistence farming and livestock herding are main sources of domestic and local economy. Rangelands being the largest land use (>33%, n = 72,496 km2) directly contribute about 38–40% of the total household income in rural communities. Apart from this, villagers have also been benefiting tremendously through trophy hunting of mountain ungulates. Uncontrolled grazing of pastures by livestock coupled with illicit removal of highly sparse and scattered natural vegetation for domestic energy have appeared affecting ecological health and productivity of highland pastures, and therefore negatively impact dependent populations of wild and domestic herbivores in the area, which may ultimately affect rural livelihood and domestic economy in the long run (Khan, 2013), However, the drivers of rangeland degradation/browning trend are yet not well understood. Therefore a detailed investigation into rangelands resource dynamics, existing and emerging pressures and the changing climatic conditions, is required for sustainable management of rangeland ecosystem and its associated biodiversity in the area.
Acknowledgements
Authors wish to thank staffs of World Wide Fund for Nature (WWF) Pakistan, Gilgit office, International Centre for Integrated Mountain Development (ICIMOD), Gilgit-Baltistan Parks & Wildlife Department, Directorate of Central Karakoram National Park and the local communities for their assistance, support and hospitality during the field study. We are also thankful to unknown reviewers for improving the draft manuscript.
References
- Prediction of Kashmir markhor habitat suitability in Chitral Gol National Park, Pakistan. Biodiversity. 2012;13(2):78-87.
- [Google Scholar]
- Population size and demographic parameters of blue sheep (Pseudoisnayaur) in Dhorpatan Hunting Reserve, Nepal. Int. J. Biodivers. Conserv.. 2010;2(3):51-56.
- [Google Scholar]
- On the distribution of Punjab urial (Ovis vignei punjabiensis) Rec. Zool. Surv. Pakistan. 2004;15:1-6.
- [Google Scholar]
- Conflicts between traditional pastoralism and conservation of Himalayan ibex (Capra sibirica) in the Trans-Himalayan mountains. Anim. Conserv.. 2004;7(2):121-128.
- [Google Scholar]
- Modeling habitat suitability for moose in coastal northern Sweden: empirical vs process-oriented approaches. AMBIO: J. Hum. Environ.. 2003;32(8):549-556.
- [Google Scholar]
- IDRISI Taiga 16.03. The IDRISI Project. University’s George Perkins Marsh, Institute at Clark University; 2009.
- ArcGIS Desktop 10. Redlands, CA: Environmental Systems Research Institute; 2011.
- ERDAS Imagine 10. Atlanta, Georgia: Leica; 2010.
- Comparison of single-year and multiyear NDVI time series principal components in cold temperate biomes. IEEE Trans. Geosci. Remote Sens.. 2003;41(11):2568-2574.
- [Google Scholar]
- Modeling habitat suitability for complex species distributions by environmental-distance geometric mean. Environ. Manage.. 2003;32(5):614-623.
- [Google Scholar]
- Which is the optimal sampling strategy for habitat suitability modelling. Ecol. Model.. 2002;157(2):331-341.
- [Google Scholar]
- Modeling the dynamics and management of ibex populations in Switzerland. 1998. p. :139-144.
- [Google Scholar]
- Assessing habitat-suitability models with a virtual species. Ecol. Model.. 2001;145(2):111-121.
- [Google Scholar]
- Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecology. 2002;83(7):2027-2036.
- [Google Scholar]
- Evaluating the ability of habitat suitability models to predict species presences. Ecol. Model.. 2006;199(2):142-152.
- [Google Scholar]
- Biomapper 4.0.. Switzerland: Laboratory for Conservation Biology, Department of Ecology and Evolution, University of Lausanne; 2007. URL: http://www.unil.ch/biomapper
- Red List of Threatened Species. 2011.2. 2012.
- Rangelands of Gilgit-Baltistan and Azad Jammu & Kashmir, Pakistan – Current Status, Values and Challenges. 2013. FAO Report: 45
- Shimshal Pamir Lakes: a prospective high altitude wetlands site for transboundary collaboration between China and Pakistan. Rec. Zool. Surv. Pak.. 2012;21:1-9.
- [Google Scholar]
- Abundance, distribution and conservation of key ungulate species in Hindu Kush, Karakoram and Western Himalayan (HKH) mountain ranges of Pakistan. Int. J. Agric. Biol.. 2014;16(6):1050-1058.
- [Google Scholar]
- Sensitivity analysis of NDVI dynamic and air temperature, precipitation changes of Chinese vegetation types. Chin. J. Plant Ecol.. 2000;24(3):379-382.
- [Google Scholar]
- Biomass Assessment Study Mountain Areas Conservancy Project. Karachi: IUCN Pakistan; 2001. IUCN Report
- Discriminant analysis of the terrestrial and aquatic habitat determinants of the smooth newt (Triturus vulgaris) and the common frog (Rana temporaria) in Ireland. J. Zool.. 1998;244(01):1-6.
- [Google Scholar]
- Restoration of fragmented landscapes for the conservation of birds: a general framework and specific recommendations for urbanizing landscapes. Restor. Ecol.. 2001;9(3):280-292.
- [Google Scholar]
- Illustrated Handbook of Animal Biodiversity of Pakistan. Centre for Environmental Research and Conservation; 1998.
- Habitat selection by large herbivores in Lake Nakuru National park, Kenya. Biodivers. Conserv.. 1997;7(1):1-8.
- [Google Scholar]
- A contextual Mann–Kendall approach for the assessment of trend significance in image time series. Trans. GIS. 2011;15(5):599-611.
- [Google Scholar]
- Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol.. 2005;20(9):503-510.
- [Google Scholar]
- Can coral reefs be monitored from space? AMBIO: J. Hum. Environ.. 2003;32(8):586-593.
- [Google Scholar]
- Ecological niches of three teuthophageous odontocetes in the northwestern Mediterranean Sea. Ocean Sci.. 2008;4(1):49-59.
- [Google Scholar]
- Modelling the habitat suitability of cetaceans: example of the sperm whale in the northwestern Mediterranean Sea. Deep Sea Res. Part I: Oceanogr. Res. Pap.. 2009;56(4):648-657.
- [Google Scholar]
- Population status of wildlife in Khunjerab National Park (Pakistan) Tiger Pap.. 1990;17(4):25-28.
- [Google Scholar]
- Reading, R.P., Amgalanbaatar, S., Batbold, J., Boldgev, B. (1995). Biological assessment of three beauties of the Gobi National Conservation Park with recommendations for special zone (Core Area) locations, tourist routes, and park management. Northern rockies conservation cooperative & nature conservation International, Jackson, Wyoming. Unpublished report.
- Biological assessment of three beauties of the Gobi National Conservation Park, Mongolia. Biodivers. Conserv.. 1999;8(8):1115-1137.
- [Google Scholar]
- The Mammals of Pakistan. Cambridge Univ Press; 1977.
- Ecological niche modelling of two cryptic bat species calls for a reassessment of their conservation status. J. Appl. Ecol. 2007
- [Google Scholar]
- Mountain Monarchs. Wild Sheep and Goats of the Himalaya. University of Chicago Press; 1977.
- Status Survey and Conservation Action Plan for Caprinae. Wild Sheep and Goats and Their Relatives. Gland, Switzerland and Cambridge, UK: World Conservation Union; 1997.
- An overview of sustainability assessment methodologies. Ecol. Ind.. 2009;9(2):189-212.
- [Google Scholar]
- Remote sensing of vegetation and land-cover change in Arctic Tundra Ecosystems. Remote Sens. Environ.. 2004;89(3):281-308.
- [Google Scholar]
- Predicting the potential distribution of invasive ring-necked parakeets Psittacula krameri in northern Belgium using an ecological niche modelling approach. Biol. Invasions 2008 on-line
- [Google Scholar]
- A presence-only habitat suitability model for large grazing African ungulates and its utility for wildlife management. Afr. J. Ecol.. 2006;45:347-354.
- [Google Scholar]
- Predicting species occurrences: progress, problems, and prospects. In: Predicting Species Occurrences. Issues of Accuracy and Scale. 2002. p. :739-749.
- [Google Scholar]




