

Translate this page into:
Exploring the efficiency of Ethyl Methane Sulfonate (EMS) and Sodium Azide (SA) to induce mutation in chili (Capsicum annuum L.) germplasm
⁎Corresponding author at: Department of Plant Breeding & Genetics, Ghazi University, Dera Ghazi Khan 32200, Pakistan. smahpara@gudgk.edu.pk (Shahzadi Mahpara)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Background
Exploring genetic diversity in traits of interest and utilizing it to breed high yielding, disease-resistant and high market value varieties is the prime objective of plant breeders. Unfortunately, genetic diversity in most of the crops has declined due to selection and reutilization of already adapted germplasm. Chemical mutagens are widely used to create novel changes in germplasm; however, these have been rarely tested for chili germplasm in Pakistan.
Methods
We used ethyl methane sulfonate (EMS) and sodium azide (SA) to induce mutations for obtaining novel alterations in morphological and yield-related traits of four chili genotypes (i.e., ‘Mexi’, ‘Dandi Cut’, ‘Nageena’ and ‘Talhari’). Firstly, concentrations of both mutagens were calibrated, i.e., 1.5 % EMS and 0.5 % SA. Afterwards, seeds of four chili cultivars were treated and grown in peat trays. As the nursery was established, seedlings were transferred to media (sand: silt: clay + 1 kg well-decomposed FYM) after 30 days of germination. Different mutants were isolated based on differential morphological and phenotypic expressions.
Results
It was found that efficiency of EMS was greater than SA and more variants were noticed in case of EMS-treated populations. Genetic analysis showed that almost all the traits were affected by mutagens in the M1 generations although the stability of isolated mutants was not checked in later generations. All the traits showed higher estimates of heritability but in some traits genetic advance was non-significant.
Conclusion
It is recommended that isolated mutants could be used as rich genetic diversity source to breed new varieties. Furthermore, EMS proved a suitable mutagen for targeted mutations.
Keywords
Capsicum
Chemical mutagen
Correlation
Genetic variability
Yield

1 Introduction
Chili, also known as Capsicum annuum L., is an essential crop for the culinary industry. Capsaicinoids, carotenoid pigments, and vitamins A and C are the main constituents of chili. Capsicum is a type of basic spice used in every dish due to its sharpness, flavor, aroma, and color. It has miraculous properties and used as anti-inflammatory, analgesic, carminative, and antioxidant (Sim and Sil, 2008). Increased yield, nutritional value, and secondary metabolite content are the main targets of contemporary breeding approaches since they all have a direct impact on crop profitability (Bosland et al., 2012; Lee et al., 2016).
Breeding in chili is mainly focused on improving economically important traits, including yield, nutrient and secondary metabolite contents (Solomon et al., 2019). Chili is a high economic value crop and has potential to revolutionize the farming community if consumers-oriented traits are incorporated in modern cultivars. The availability of genetic diversity is indispensable for such breeding programs which can be achieved by creating novel alleles through altering nucleotide sequences by targeted mutations (Arisha et al., 2015; Hwang et al., 2014). Chemical mutagens cause morphological mutations that are expressed in first mutagenic generation and scientists have successfully isolated morphological mutants from chili (Manzila and Priyatno, 2020; Muslihatin, 2020).
Variability is the starting point for plant breeders and mutation breeding is an efficient and effective tool for the creation of novel variants (Hussain et al., 2021b, 2021a). Conventional breeding should be equipped with novel techniques for the alteration of genes at genic level which is depicted in phenotypic expression (Hussain et al., 2018). Chances of success increase as the spectrum of variation broadens and availability of reliable variants ease the selection procedures (Dhumal and Bolbhat, 2012). It is evident from the reported literature that chemically induced mutations of seed and other plant parts have rewarded successful alteration in genetic makeup of plants (Devi and Mullainathan, 2012). Selection of chemical mutagen is indispensable to produce high frequency of desirable mutation in any mutation breeding plan (Roychowdhury and Tah, 2011).
Although dominant mutations are often easy to see in the M1 generation, most induced mutations are recessive in nature and manifest themselves with various symptoms in the M2 generation. It is unclear how effective M1 is as a mutagen, although the presence of phenotypes such as chlorophyll deficits (such as albino plants), pollen sterility, stunting, curled leaves, and premature or delayed flowering raises suspicions (Alcantara et al., 1996; Sega, 1984; Shah et al., 2015). Recessive mutations can be observed by visual inspection and characterization of mutant phenotypes at those early stages. Morphological mutations are easy to detect if desirable characteristics are set as a criterion and used as markers to isolate candidate mutants. Plant scientist could use plant height, fruit color, disease and pest resistance, or flowering time and length of stigma or style as morphological markers for the early detection of desirable mutants (Raghuvanshi and Singh, 1982; Talebi et al., 2012).
Genetic variability is the prerequisite of a successful breeding program; however, genetic base of almost all the crops has narrowed down due to continuous selection and reutilization of already adapted genotypes. To broaden genetic base and create novel diversity, mutations are an important tool to alter the sequence of nucleotides, which may lead to the formation of new alleles useful for crop improvement (Dhumal and Bolbhat, 2012; Restaino, 1983).
For this reason, the present experiment was designed to induce chemical mutagenesis in chili and create genetic variability required for crop improvement programs. It is predicted that mutations broaden the genetic base and provide a wide spectrum of genotypic as well as phenotypic variability. For variability, we used morphological characteristics of mutants because phenotypic expression is a robust and reliable method to detect interspecific genetic variability. It was hypothesized that different mutagens used in the study would induce significant mutations in chili germplasm which would help in the selection of traits of interest.
2 Materials and methods
2.1 Experimental site
The experiment was conducted at Plant Breeding and Genetics Department, Ghazi Univeristy, Dera Ghazi Khan, Pakistan during 2019–2020.
2.2 Experiment details
Four chili genotypes, i.e., ‘Mexi’, ‘Nageena’, ‘Dandi cut’ and ‘Talhari’ were treated with chemical mutagen, i.e., 0.5 % sodium azide (SA) and 1.5 % ethyl methane sulphonate (EMS). Stock solutions of SA and EMS were prepared and then working solutions were made on percent basis. The seeds were surface disinfected and then washed under tap water for 1 h. After disinfecting, seeds were soaked in tap water for 3 h and then presoaked seeds were treated with 1.5 % EMS and 0.5 % SA solution. After carefully removing seeds from mutagen solution, these were treated with 0.1 M sodium phosphate buffer (pH 7.0) to prevent rapid hydrolysis. Finally, seeds were washed under running tap water for 1 h. Control seeds were treated with 0.1 M phosphate buffer (pH 7.0) and then preserved. All the precautionary measures were strictly followed. Treated seeds were sown in pots having sand: silt: clay (1:1:1) along with well decomposed farmyard manure. All the plant production and protection practices were followed as per rule.
2.3 Data collection
Data relating to fruit width, internodal length, fruit length, seed index, peduncle length, fresh fruit weight, plant height, dry fruit weight, number of primary branches and number of seeds per fruit were collected at maturity. Five randomly selected plants were used from each genotype and mutagen to collect the data. Fruit length and width, and internodal length were measured with the help of a vernier caliper. Five fruits were randomly used for the measurements. A meter rod was used to measure the heights of five randomly selected plants and then averaged. The number of primary branches were counted from five randomly selected plants and averaged. The number of seeds present in five randomly selected seeds from each treatment were counted and averaged.
2.4 Experimental setup
The experiment was set up using a randomized full block design with split plots. Genotypes of peppers were maintained in major plots, whereas mutations were randomly distributed across smaller plots. Three separate containers were used to cultivate the seeds, and the results from these containers were a replication. There were 4 replicates of each treatment. Upon reaching harvest maturity, chili plants were removed from the experiment and information on the many qualities mentioned above was gathered.
2.5 Statistical analysis
Collected data on different morphological and yield-related traits were by two-way analysis of variance (ANOVA) to test the significance. Differences among treatments’ means were determined by Tukey’s post-hoc test (Steel et al., 1997). Heritable variability for different attributes was estimated. Correlation was figured out following procedures of (Dewey and Lu, 1959). All analysis were done on SPSS statistical software version 21.0.
3 Results
3.1 Morphological and yield-related traits
With a few notable exceptions, analysis of variance revealed that both the additive and multiplicative impacts of mutagens and varieties significantly affected all attributes under study (Table 1). In this work, mutagenic treatments were used to enhance growth and yield-related parameters of chili genotypes (Fig. 1). FW = fruit width, INL = internodal length, FL = fruit length, SI = seed index, PL = peduncle length, FFW = fresh fruit weight, PH = plant height, FDW = dry fruit weight, PB = number of primary branches, SF = number of seeds per fruit. * and ** indicate that the relevant trait was significantly altered by the individual and interactive effects of mutagens or varieties, whereas ns indicates that the relevant treatment was not significantly affected by the individual and interactive effects of mutagens or varieties.
SOV
FW
FL
PL
NPB
INL
PH
SI
FFW
DFW
SPF
Treatment (T)
0.10312*
0.29558 *
0.00134ns
0.09784*
0.06861*
20.7601**
0.06457*
0.01479*
0.01448*
9.3825**
Varieties (V)
1.42252*
1.26968**
0.00303*
0.16122*
0.48186*
15.8320**
0.03768*
0.19378*
0.06628*
44.1270**
T × V
0.23008**
0.18097*
0.00607**
0.38346*
0.08068*
4.2988*
0.02268*
0.01177*
0.00128ns
14.8844*
Error
0.36153
0.24172
0.00785
0.23931
0.04982
6.4769
0.03356
0.02241
0.03360
26.4231

The EMS and SA-treated plants showing phenotypic variability.
The highest fruit width (7.067) was noted for the genotype ‘Dandi Cut’ treated with 1.5 % EMS followed by ‘Nageena’ variety treated with 0.5 % SA. The lowest fruit width was recorded for ‘Talhari’ genotype treated with buffer solution only. The genotype ‘Dandi Cut’ treated with 0.5 % SA recorded the highest fruit length (8.71) by ‘Mexi’, ‘Talhari’ and ‘Nageena’ varieties treated with 0.5 % SA. The lowest fruit length was recorded for ‘Mexi’ variety treated with none of the mutagens included in the study. Genotype ‘Mexi’ observed the highest peduncle length (2.27) in when treated with 1.5 % EMS followed by ‘Dandi Cut’, ‘Talhari’ and ‘Nageena’ genotypes treated with 1.5 % EMS (Fig. 2).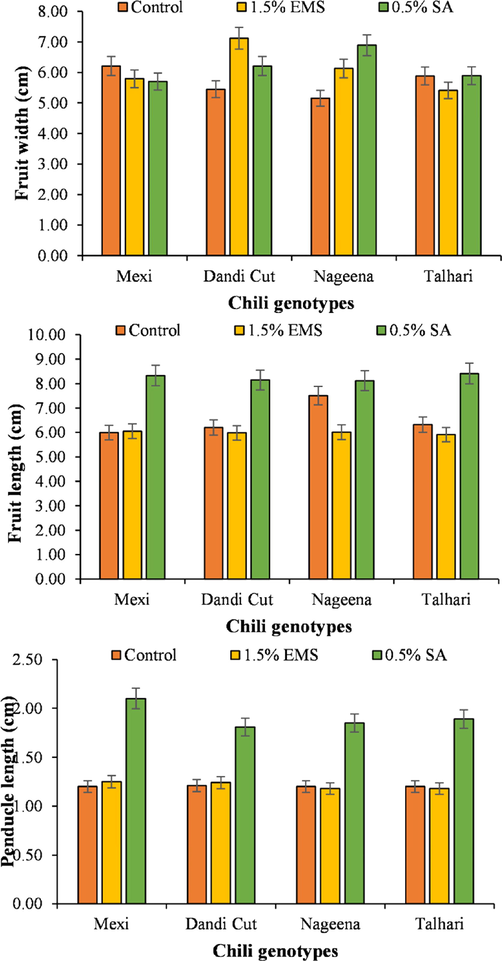
Fruit width, fruit length and peduncle length of chili genotypes treated with SA and EMS.
Genotype ‘Mexi’ produced higher number of primary branches (6.58) without application of any mutagen followed by mutation-induced ‘Mexi’ with 1.5 % EMS. The lowest number of primary branches were noted for varieties ‘Dandi Cut’ and ‘Talhari’ treated with 1.5 % EMS. The genotype ‘Mexi’ recorded the highest internodal length (6.3) in no mutagen application treatment followed by mutation-induced ‘Dandi Cut’ with 1.5 % EMS and ‘Mexi’ treated with 0.5 % SA. The lowest internode length was observed for variety ‘Nageena’ receiving no mutagen treatment. The variety ‘Dandi Cut’ resulted in the highest plant height (68.15) when treated with 1.5 % EMS followed by ‘Mexi’ (67.5), ‘Talhari’ (67.55) and ‘Nageena’ experiencing treatment with 1.5 % EMS (Fig. 3).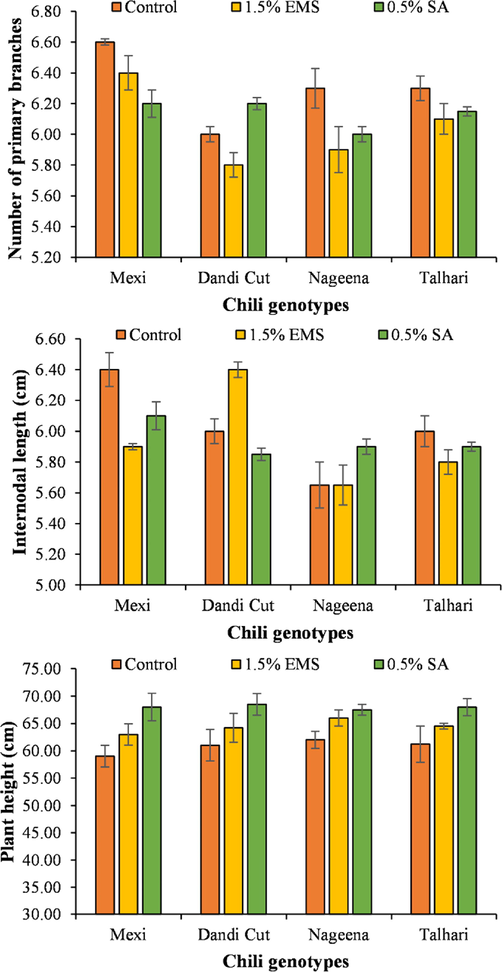
Number of primary branches, internode length and plant height of chili genotypes treated with SA and EMS.
The highest seed index (3.18) was observed for the variety ‘Dandi Cut’ treated with 1.5 % EMS followed by variety ‘Mexi’ treated with 1.5 % EMS and 0.5 % SA, respectively. The lowest seed index was recorded for the genotype ‘Nageena’ receiving no application of mutagens and ‘Dandi Cut’ treated with 1.5 % EMS. The highest fresh fruit weight (1.66) was recorded for mutation-induced ‘Mexi’ genotype treated with 0.5 % SA followed by ‘Dandi Cut’ and ‘Mexi’ receiving no mutagen application, ‘Mexi’ treated with 1.5 % EMS and ‘Talhari’ receiving 0.5 % SA. The genotype ‘Mexi’ recorded the highest fruit dry weight (1.53) under 1.5 % EMS application followed by ‘Mexi’ with no mutagen application and 0.5 % SA applicaiton, ‘Talhari’, and ‘Nageena’ with 1.5 % EMS. The lowest fruit dry weight was recorded for the genotype ‘Dandi Cut’ receiving 1.5 % EMS and no mutagen application (Fig. 4).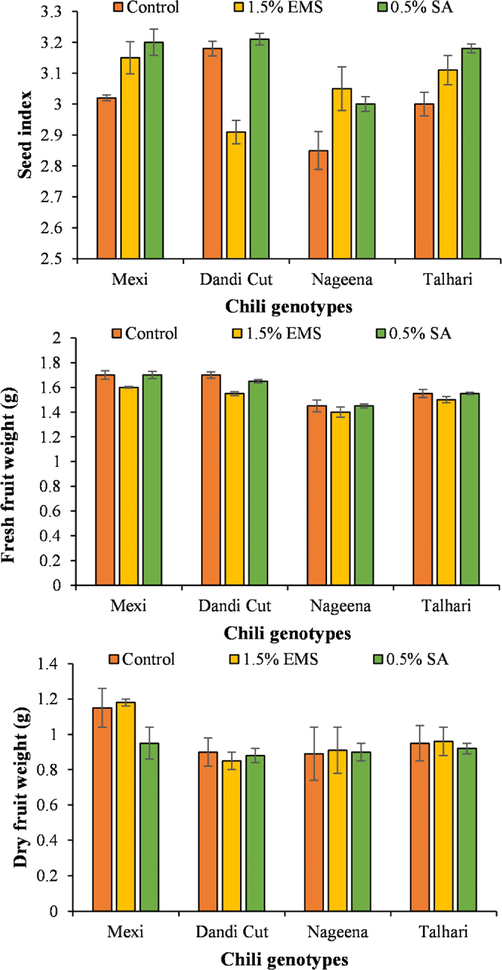
Seed index, fresh fruit weight and dry fruit weight of four genotypes treated with SA and EMS.
The genotype ‘Mexi’ resulted in the highest number of seeds per fruit (94.33) in mutagen treatment 0.5 % SA followed by variety ‘Talhari’, ‘Dandi Cut’ and ‘Nageena’ receiving 0.5 % SA. The lowest number of seeds per fruit were noted for the genotype ‘Dandi Cut’ experiencing no mutagen application (Fig. 5).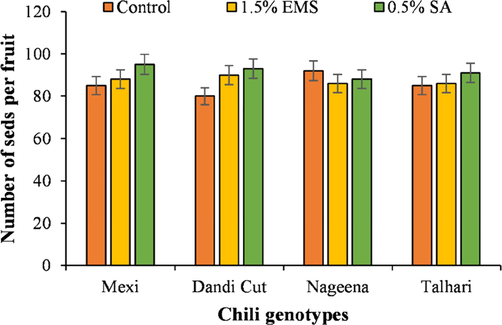
Number of seeds per fruit of chili genotypes treated with SA and EMS.
3.2 Genetic variability, heritability, and genetic advance of mutant population
In contrast to the untreated control, all four types of the mutant population (M1) were tested for phenotypic and genotypic diversity, heritability, genetic advancement, and association. Fruit width exhibited substantial heritability (99.8 %), significant genetic advancement (71.0 %), and 15.5 and 15.5 genotypic and phenotypic variance, according to the results. Fruit length showed moderate genetic advancement of 47.0 %, genotypic and phenotypic variability of 1.83 and 1.85, and heritability of 99.3 %. With a high heritability (91.9 %) and a low genetic advance (32.6 %), the genotypic and phenotypic variances for peduncle length were 0.22 and 0.23, respectively (Table 2). FW = fruit width, INL = internodal length, FL = fruit length, SI = seed index, PL = peduncle length, FFW = fresh fruit weight, PH = plant height, FDW = dry fruit weight, PB = number of primary branches, SF = number of seeds per fruit.
Variable
VG
VP
h2bs (%)
GCV
PCV
GA
GA%
FW
15.5007
15.5277
99.84
0.4742
8.7312
0.5974
71.077
FL
1.8322
1.8556
99.38
0.3582
4.1372
0.4814
47.061
PL
0.2214
0.23
91.91
0.3012
0.7422
0.4244
32.682
PB
0.1402
0.3396
79.37
0.3952
2.6172
0.5184
43.500
INL
1.0333
1.0414
99.10
0.2722
2.0792
0.3954
29.371
PH
198.7148
199.7441
99.48
0.3352
29.2322
0.4584
42.488
SI
0.0403
0.033
99.62
0.4392
2.9922
0.5624
63.895
FFW
2.9638
2.9657
99.95
0.7232
4.7062
0.8464
122.547
DFW
0.2505
0.256
95.86
0.4182
0.8432
0.5414
57.299
SPF
278.9415
280.133
99.57
0.3422
34.5782
0.4654
43.802
The main branches had genetic progress (43.5 %), moderate heritability (79.7 %), and genotypic and phenotypic variations of 0.14 and 0.33. Internode length had 99.1 % heritability, 29.3 % genetic progress, and genotypic and phenotypic variances of 1.03 and 1.04. Plant height had high heritability (99.5 %) and modest genetic progress (42.4 %). The seed index had genotypic and phenotypic variances of 0.04 and 0.03, high heritability (99.6 %), and moderate genetic advancement (63.8 %).
Genotypic and phenotypic variance were 2.96 and 2.98 for fresh fruit, respectively, with high heritability (99.9 %) and rapid genetic progress (122.5 %), whereas these values were 0.25 and 0.26 for dry fruit, with high heritability (99.6 %) and moderate genetic progress (57.2 %). High heritability (99.5 %) and modest genetic progress (43.802 %) were found in the seeds produced per fruit, with 278.94 and 280.133 variation in genotype and phenotype, respectively (Table 2).
3.3 Genotypic correlation among growth and yield contributing traits of mutant population
Fruit width and fruit fresh weight were positively correlated, however fruit width and either the seed index or fruit length were negatively correlated. The height of the plant, the length of the internodes, and the length of the fruit were all significantly related to one another. The relationship between peduncle length and the separation between nodes was statistically significant. Primary branch length and internodal length were negatively correlated. Although there was a negative and statistically significant link with main branch count, there was a positive relationship with inter nodal length, fruit length, and peduncle length. The size of the fruit correlated with plant maturity. The fruit's diameter and the seed index were strongly negatively correlated. The weights of fresh and dried fruits showed a very strong positive correlation. A fruit's “seed index” and its seed content are positively correlated (Table 3). FW = fruit width, INL = internodal length, FL = fruit length, SI = seed index, PL = peduncle length, FFW = fresh fruit weight, PH = plant height, FDW = dry fruit weight, PB = number of primary branches, SF = number of seeds per fruit. * and ** indicate that the relevant traits had significant positive or negative correlations.
DRFW
FL
FRFW
FW
INL
PB
PH
PL
SI
FL
0.0324
FRFW
0.5329**
−0.0321
FW
0.3425
−0.5354*
0.5923**
INL
0.2182
0.5921**
0.1163
0.2268
PB
−0.0234
−0.0011
−0.0587
−0.227
−0.564**
PH
0.1233
0.5341**
0.1121
0.1762
0.2123
0.1235
PL
−0.2341
−0.1121
−0.0812
−0.172
0.5352**
0.1029
−0.123
SI
0.1023
0.0526
−0.1672
−0.59*
−0.0011
−0.152
−0.091
0.0212
SPF
0.0231
−0.0211
0.0213
0.2213
−0.0123
−0.213
0.0013
−0.322
0.5893**
4 Discussion
In addition to revealing substantial genotypic differences, the data demonstrated how mutagenetic treatments influenced plant development and productivity in unexpected ways. These results imply that chemical mutagens either boosted or drastically increased cell proliferation, which in turn enhanced or lowered fruit traits. Mutagenic agents may increase genetic diversity in mutant offspring. However, this method has been used to improve multiple improved varieties in a number of cultural contexts, so it must have some positive effects (Ando and Montalván, 2001). Wheat, barley, rice, cow beans, cotton, and peanuts, all of which are seed-propagated, have all benefited from intentional mutation in order to be made more nutritious (Khan et al., 2009). Chromosome mutations could be a factor in the emergence of mutant plants. These mutations may change an organism's whole genome or the number of chromosomes in it, among other things (Jo et al., 2016).
Both sodium azide (SA) and ethyl methane sulphonate (EMS) are efficient mutagens for introducing genetic variation into plant populations. In this work, 1.5 % EMS and 0.5 % SA were applied to four chili genotypes, and mutant populations were identified based on variations in yield-related characteristics and phenotypic expressions. The results showed that mutagens affected almost every attribute linked to growth and yield, but the most significant impacts were shown in qualitative variables due to obvious phenotypic expression. The findings of the first discovered qualitative variations were positive, and several new mutants were identified. It is commonly known that a breeding program needs genetic variety to be effective. Therefore, it's crucial to increase the effectiveness and efficiency of the mutagens utilized to produce this variety (Chattopadhyay et al., 2011; Devi and Mullainathan, 2012).
Chemical mutagens may successfully alter a plant's genetic makeup, albeit the degree of success is highly dependent on the quantity of the chemical and the kind of plant used in the experiment. Since a high mutagen concentration prevents seed germination due to loss of viability, breeders must identify the proper mutagen concentration without compromising seed viability (Ando and Montalván, 2001; Khan et al., 2009). Some chemical mutagens have been shown to inhibit seed germination at high doses and, more crucially, to disrupt ecosystems at these levels (Jo et al., 2016). To that end, we found the best balance between SA and EMS in the current investigation.
Due to the recessive nature of the majority of induced mutations, only dominant mutations may be identified, and mutant phenotypes are unlikely to appear in the M1 generation. It is claimed that chemical mutagens cause visible morphological changes that might act as signal characteristics for spotting mutations (Hussain et al., 2021b, 2021a; Jyothi et al., 2011; Talebi et al., 2012). We found a number of mutant traits in M1 plants, including as stunted growth, abnormal stamen and pistil formation, sterility in the pollen, and uneven stem and leaf development.
Microsporogenesis is severely impaired by several mutations, leading to the generation of infertile gametes (Kumar et al., 2007). The results of previous mutation experiments in pepper and eggplant have similarly pointed to the occurrence of mutant traits in M1 plants (Ando and Montalván, 2001), even if environmental and managerial causes may be at play in certain cases (Hussain et al., 2018). Due to differing estimations of heritability and genetic progress, the mutation frequencies seen in generation M1 may vary substantially in generations M2 and M3 (Shah et al., 2015).
The leaf morphology of mutant plants was discovered to be odd, with long petioles, rough leaves, or a sprouting and bunchy appearance. Additionally, five mutants exhibiting signs of viral infection, such as leaves that folded up or down, were observed, but no viruses were discovered. Curled leaves are a consequence of a helpful mutation that speeds up photosynthesis by boosting how much light they can absorb. Additionally, it could change leaf transpiration and increase resilience to drought. As a consequence, genetic resources that might be employed to create crops resistant to drought are drawn to wrinkled leaf variations (Bosland et al., 2012). Notably, independent of humidity and temperature, the curling trait persisted during the whole development period (Lee et al., 2016).
The market value of a horticulture product might rise or fall depending on whether or not consumers prefer a certain fruit shape. It's possible that new varieties of fruit might be grown from mutants with unusual forms to meet rising consumer demand. The present study was able to successfully isolate mutants with round, semi-round, and pyramid-shaped fruits, despite the complexity and difficulty of evaluating fruit form. Perhaps in the not-so-distant future, these variants will be used in breeding initiatives. There is also a lack of understanding about the mechanisms that determine pepper fruit size, shape, and orientation (Oleykowski et al., 1998). The aforementioned mutations will be used in genetic research to provide information on the physiological processes impacted by altering the genetic makeup of plants that control aspects related to fruit shape. The pepper fruit's economic value is significantly influenced by its hue. We found a lineage that produced fruit that was consistently orange, and a recent study utilized this mutant to look at the inheritance patterns related to fruit color in peppers (Dhumal and Bolbhat, 2012). Therefore, the mutations found here may function as a beginning point for locating and describing the genes that control fruit form and color.
The coefficients for phenotype were higher than those for genotype because it depicts the interplay between genotype and environment. Heritability estimates were generally high, which suggests that mutations may be handed on down the generations (Hosamani, 2003; Singh et al., 2017). Findings reveal the mutant population has desirable diversity and heredity, suggesting it might be evaluated in a variety of environments as part of chili improvement initiatives. The previous investigations all seemed to arrive at similar conclusions. High-heritability, high-genetic-progress mutants could be useful in chili breeding projects (Das and Maurya, 2004; Mini and Khader, 2004; Singh et al., 2009). The examination of correlation showed that there is a weak but negative association between fruit width and seed index, and that there is a weak but positive relationship between fruit width, internode length, plant height, and the number of seeds found in each fruit. Numerous previous studies found a significant correlation between the height of the plant, the length of the internodes, and the size of the fruit (Kumar et al., 2003; Sreelathakumary and Rajamony, 2006). There is a substantial association between the length of the peduncle and the distance between the nodes. Researchers performed experiments on chili like those that came before them, and they found comparable findings (Chattopadhyay et al., 2011; Jyothi et al., 2011; Mishra et al., 1998).
5 Conclusion
We used EMS and SA in the present testing to create distinctive variations in advanced chili cultivars. We found that the EMS mutagen was more effective than SA. When the concentration of SA was increased from 0.5 % to 1.5 % during the calibration of mutagens, seed viability started to decline, but EMS had no impact on germination at that concentration. It was revealed that the EMS mutagen significantly altered the morphological and yield characteristics in the M1 generation. The mutations that were discovered during this study might be useful in several chili breeding projects. It is advisable to use an EMS mutagen in targeted mutagenesis, and changes should be studied at the molecular level.
Acknowledgement
This project was supported by Researchers Supporting Project number (RSP2023R5) King Saud University, Riyadh, Saudi Arabia. This research was also funded by RMC-UTM through the industrial projects Nos. RJ130000.7609.4C465 and RJ130000.7609.4C359.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Ethyl methanesulfonate-induced seed mutagenesis of Capsicum annuum. J. Hered.. 1996;87:239-241.
- [Google Scholar]
- Gamma-ray radiation and sodium azide (NaN3) mutagenic efficiency in rice. Crop. Breed. Appl. Biotechnol.. 2001;1
- [Google Scholar]
- Ethyl methane sulfonate induced mutations in M2 generation and physiological variations in M1 generation of peppers (Capsicum annuum L.) Front. Plant Sci.. 2015;6:399.
- [Google Scholar]
- Diversity of genetic resources and genetic association analyses of green and dry chillies of Eastern India. Chil. J. Agric. Res.. 2011;71:350.
- [Google Scholar]
- Effect of gamma rays and ethyl methane sulphonate (EMS) in M 3 generation of blackgram (Vigna mungo L. Hepper) African J. Biotechnol.. 2012;11:3548-3552.
- [Google Scholar]
- A Correlation and Path-Coefficient Analysis of Components of Crested Wheatgrass Seed Production 1. Agron. J.. 1959;51:515-518.
- [Google Scholar]
- Induction of genetic variability with gamma radiation and its applications in improvement of horsegram. Gamma Radiat. 2012:207-228.
- [Google Scholar]
- Hosamani, R.M., 2003. Variability, correlation and path analysis in kharif-grown chilli (Capsicum annuum L.) genotypes for different characters.
- Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manage. 2018
- [CrossRef] [Google Scholar]
- CaWRKY30 Positively Regulates Pepper Immunity by Targeting CaWRKY40 against Ralstonia solanacearum Inoculation through Modulating Defense-Related Genes. Int. J. Mol. Sci.. 2021;22:12091.
- [Google Scholar]
- A basic helix-loop-helix transcription factor CabHLH113 positively regulate pepper immunity against Ralstonia solanacearum. Microb. Pathog. 2021
- [CrossRef] [Google Scholar]
- Phenotypic variants among ethyl methanesulfonate M2 mutant lines in Capsicum annuum. Plant Genet. Resour.. 2014;12:S141.
- [Google Scholar]
- Construction of mutation populations by gamma-ray and carbon beam irradiation in chili pepper (Capsicum annuum L.) Hortic. Environ. Biotechnol.. 2016;57:606-614.
- [Google Scholar]
- Variability studies in chilli (Capsicum annuum L.) with reference to yield attributes. J. Hortic. Sci.. 2011;6:133-135.
- [Google Scholar]
- Sodium azide: a chemical mutagen for enhancement of agronomic traits of crop plants. Environ. We Int. J. Sci. Tech. 2009;4:1-21.
- [Google Scholar]
- Kumar, B.K., Munshi, A.D., Joshi, S., Kaur, C., 2003. Correlation and path coefficient analysis for yield and biochemical characters in chilli (Capsicum annuum L.).
- Hexavalent chromium removal from wastewater using aniline formaldehyde condensate coated silica gel. J. Hazard. Mater.. 2007;143:24-32.
- [Google Scholar]
- Genetic diversity and population structure analysis to construct a core collection from a large Capsicum germplasm. BMC Genet.. 2016;17:1-13.
- [Google Scholar]
- Manzila, I., Priyatno, T.P., 2020. Genetic variations of EMS-induced chili peppers (Capsicum annuum) cv. Gelora generate geminivirus resistant mutant lines. In: IOP Conference Series: Earth and Environmental Science. IOP Publishing, pp. 12031.
- Mini, S., Khader, K.M.A., 2004. Variability, heritability and genetic advance in wax type chilli (Capsicum annuum L.).
- Correlation and path analysis in sweet pepper (Capsicum annuumL.) Recent Hort. 1998;4:123-126.
- [Google Scholar]
- Effect of chemical mutagen EMS (Ethyl Methane Sulfonate) on growth and phytochemical response of Bara chilli variety (Capsicum frutescens var. bara) Ann. Biol.. 2020;36:186-189.
- [Google Scholar]
- Mutation detection using a novel plant endonuclease. Nucleic Acids Res.. 1998;26:4597-4602.
- [Google Scholar]
- Induced mutagenesis in capsicum L. II. Effects of single and combined mutagenic treatments on habit and fruit. Curr. Sci.. 1982;51:235-237.
- [Google Scholar]
- Restaino, F., 1983. [Pepper brachytic forms (Capsicum annuum L.) obtained through induced mutagenesis].[English); It (Italian]. Genet. Agrar.
- Genetic variability study for yield and associated quantitative characters in mutant genotypes of Dianthus caryophyllus L. Int. J. Biosci. 2011;1:38-44.
- [Google Scholar]
- A review of the genetic effects of ethyl methanesulfonate. Mutat. Res. Genet. Toxicol.. 1984;134:113-142.
- [Google Scholar]
- Effect of ethyl methyl sulfonate concentration and different treatment conditions on germination and seedling growth of the cucumber cultivar Chinese long (9930) Genet. Mol. Res.. 2015;14:2440-2449.
- [Google Scholar]
- Antioxidant activities of red pepper (Capsicum annuum) pericarp and seed extracts. Int. J. food Sci. Technol.. 2008;43:1813-1823.
- [Google Scholar]
- Genetic variability, heritability and genetic advance in chilli (Capsicum annuum L.) genotypes. Int. J. Curr. Microbiol. App. Sci. 2017;6:2704-2709.
- [Google Scholar]
- Genetic variation, association of characters, and their direct and indirect contributions for improvement in chilli peppers. Int. J. Veg. Sci.. 2009;15:340-368.
- [Google Scholar]
- Genetic diversity and population structure of Ethiopian Capsicum germplasms. PLoS One. 2019;14:e0216886.
- [Google Scholar]
- Variability, heritability and genetic advance in chilli (Capsicum annuum L.) J. Trop. Agric.. 2006;42:35-37.
- [Google Scholar]
- Steel, R., Torrei, J., Dickey, D., 1997. Principles and Procedures of Statistics A Biometrical Approach., A Biometrical Approach.
- Talebi, Ali Benjavad, Talebi, Amin Benjavad, Shahrokhifar, B., 2012. Ethyl methane sulphonate (EMS) induced mutagenesis in Malaysian rice (cv. MR219) for lethal dose determination.




