

Translate this page into:
Exogenous application of melatonin mitigate the heat stress in different tomato (Solanum lycopersicum L.) cultivars
⁎Corresponding author. ayazleo46@yahoo.com (Syed Ayyaz Javed)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
Heat stress alleviates yield and production of horticultural crops and is considered a major risk for sustainable agriculture. Melatonin acts as a first line soldier to regulate plant growth by creating a conducive environment under heat stress. Thus, the present study was performed in pots to mitigate the adverse impact of heat stress on four genotypes of tomato (two were heat sensitive and two were heat tolerant) through exogenous application of melatonin. The experiment was comprised of two treatments, control and foliar spray of melatonin @ 25 µM, under factorial design (CRD) with five replications by using heat-sensitive and heat-tolerant cultivars of tomato as a test crop. All the tomato varieties were treated with 25 µM melatonin through foliar spray and compared with control which remained untreated. All the growth and yield parameters and enzymatic activities of antioxidants including catalase (CAT), superoxide dismutase (SOD), and peroxidase (POD) were increased significantly due to the application of melatonin. Moreover, the melatonin spray enhanced the osmolyte production such as malondialdehyde (MDA), proline contents, total soluble sugars, and fruit quality over control treatment. Concludingly, the heat-tolerant cultivars performed better under melatonin spray as compared to heat-sensitive cultivars of tomato.
Keywords
Antioxidant enzymes
Heat stress
Melatonin
pH of tomato
Reactive oxygen species
Total soluble sugars

1 Introduction
Global warming is a serious cause of climate change owing to the increase in temperature of the universe. Thus, climate change involves in prompting various types of abiotic stresses including drought, salinity, heavy metal, and heat. Stress caused by heat is particularly a major threat for crop production including cereal crops and horticultural crops in a wide range of the world (Ohama et al., 2017) primarily in areas of low rainfall and high temperature. Heat stress disrupts the cellular and molecular mechanism of plants leading to plant wilt. Water scarcity faced by plants under heat stress leads to disorganized ion homeostasis mechanism, photosynthetic process, abated membrane stability index, increased malondialdehyde (MDA) contents and reactive oxygen species (ROS), misfolding of protein structure due to the hampered synthesis of protein and reduced cell division (Ahmad et al., 2019). The plant is known as a sessile organism, thus under stress conditions plant triggers defensive mechanisms against unfavorable environment by scavenging ROS species to reduce oxidative stress (Mittler et al., 2011). The antioxidant enzymes including SOD, POD, CAT, and APX, and non-enzymatic antioxidants such as glutathione (GHS), phenols, and carotenoids (Jahan et al., 2019) control a defensive system within plants for its protection against stressful environment. However, heat stress is a vital issue for growing horticultural crops and food security in the future for the increasing population around the world.
Tomato (Solanum lycopersicum L.) belongs to the Solanaceae family (Aldubai et al., 2022) and is native to Peru and Mexico (Guo et al., 2022). It has substantial importance in horticultural crops due to its significant role in economic and nutritional values. Tomato and its byproducts are full of nutrition including vitamins, proteins, and antioxidants which reduce health risks in human beings, especially for cancer and heart-related individuals (Siddiqui et al. 2019). The tolerance level of abiotic stresses varies from plant to plant and species to species. Thus, heat stress is also a key factor in stunted development and reduced productivity of tomato plants. Tomato plants induce numerous natural phytohormones as brassinosteroids (BRs), ethylene, auxins (Bouzroud et al., 2018), abscisic acid, and melatonin (Altaf et al., 2021) to cope with stressful environment (Altaf et al., 2022).
Melatonin is a hormone or molecule induced naturally in both animals and plants (Kaya et al., 2019; Siddiqui et al., 2020). It plays very significant biological functions in plants against several types of stress environments by prompting a strong defensive mechanism as a master regulator in plants (Kaya et al., 2020). Melatonin not only copes with heat stress but also ameliorates root architecture, seed germination, and vigorous plant growth. It is also capable of regulating nitrogen metabolism (Zhang et al., 2017) and gene expression linked with physiological traits of plants (Arnao and Hernández-Ruiz (2019)).
Melatonin is a multi-signaling plant molecule found as a bio-stimulant and master growth regulator to promote leaf area and photosynthesis process by scavenging ROS species. Reactive oxytocin is increased directly by the system. Reactive nitrogen strains (RNS) are also produced under abiotic and biotic stress environment. The foliar melatonin spray on plants promotes their growth by improving antioxidant activities, and photosynthetic pigment and ensures their heat, drought, salt, pathogenic, heavy metal, cold stress, and acidic rain tolerances (Debnath et al., 2019). Melatonin decreases reactive oxygen species (H2O2 and O2–) levels and improves PSII activity under NaCl stress in plants (Yin et al., 2019). Furthermore, Melatonin has also the ability to improve the growth of the plant under cold stress (Bajwa et al., 2014; Turk et al., 2014). It was hypothesized that the application of melatonin can induce stress tolerance in tomato plants. Hence the current investigation was aimed to evaluate the ameliorative effect of melatonin applied through foliar spray on different cultivars of tomato grown under heat-mediated stress.
2 Materials and methods
Seeds of selected heat tolerant (T60 F1 and Super cash F1) and heat sensitive (Naqeeb and Nagina) genotypes of tomato were sanitized with a solution of 5 % sodium hypochlorite followed by repetitive cleaning with water (double distilled). Seeds were grown in plastic pots (30 cm in height and 25 cm in diameter) containing 7 kg peat moss (Sia Pendstrup Ltd., Talsii, Latvia) as a growth medium. At a suitable distance in a pot three seeds were sown and after the emergence of seeds per pot density of the plant was adjusted to one and unhealthy or less vigorous 2 seedlings were eradicated. Half-strength Hoagland solution (Table 1.1) was used as a nutrition source for plants. Irrigation was done as per the requirement of the plant by detecting the moisture level of the media. Pots were placed under the field environmental conditions at the research area of the Department of Horticulture, University of Sargodha, Sargodha, Pakistan. The average climate of Sargodha has a maximum/ minimum temperature of 41/5 °C and precipitation of 114/4 mm over a year (Fig. 1).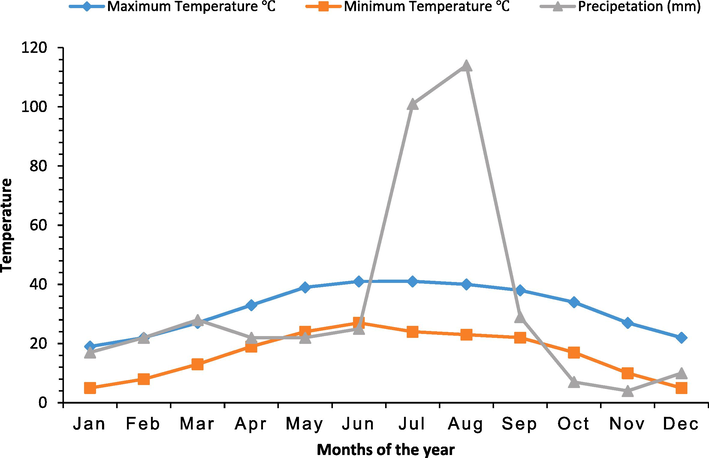
Average maximum temperature (℃), minimum temperature (℃) and precipitation (mm) of Sargodha over a year.
2.1 Treatment plan
In this experiment, 4 different genotypes (2 were susceptible and 2 were resistant to high temperature) of tomato were used with five replications and melatonin was applied through the foliar application on each cultivar in two concentrations including control as
T1 = without melatonin spray (control).
T2 = 25 µM (melatonin).
Various morpho-physiological parameters were noted by using standard methods. The experiment was performed under two factorial arrangements of a completely randomized design.
2.2 Plant attributes
Root fresh biomass and shoot fresh biomass were estimated by using a digital balance. These plants were dried in an oven (Memmert-110, Schaiwabach, Germany) at 71 °C for 2 days and dry biomasses of roots and shoots were recorded.
After 40 days of tomato seed sowing, the root and shoot length of plant samples were measured with the help of a meter rod in centimeters. Similarly, leaves and flowers per plant were counted manually.
2.3 Antioxidant enzymes
Superoxide dismutase activity was estimated from a method illustrated by Giannopolitis and Ries, 1977 by using a spectrophotometer (model M36, Beckman, CA, USA). The activity of peroxidase was determined by assessing H2O2 peroxidation with guaiacol as an electron donor (Chance and Maehly, 1955) by using a spectrophotometer (model M36, Beckman, CA, USA). Catalase activity was assayed by computing the conversion rate of hydrogen peroxide to water oxygen molecules, following the method as explained by Chance and Maehly (1955).
2.4 Malondialdehyde
Malondialdehyde (MDA), as a degradation product of lipid peroxidation, was determined by Fu & Huang (2001) with the modifications listed by Mestre et al., (2012). The MDA concentration was calculated using an extinction coefficient for MDA of 155 mm-1cm−1.
2.5 Proline
Ninhydrin, toluene, and acetic acid were used for the assessment of proline in the leaf and roots of tested tomato genotypes by using a double beam spectrophotometer (Hitachi-120, Japan), and then the toluene was taken as a blank reading according to the method described by Bates et al., (1973).
2.6 Total soluble sugars and pH
Total soluble sugar from tomato juice was determined according to the AOAC method (AOAC, 2005) by using a digital refractometer (PR-100, Atago Co. Ltd., Tokyo, Japan) and results were expressed in °Brix. In the same way, a pH meter (CH-8603, Mettler-Toledo GmbH, Schwerzenbach, Switzerland) was used to determine the acidity level of homogenized tomatoes.
2.7 Quality parameters
The number of fruits per plant was measured manually for each replication. Fruit size and its diameter were measured through a digital Vernier caliper in mm. The fruit weight of tested genotypes was measured through digital weight balance. Fruit color development was estimated manually according to the scoring method. The scores and their scale were as:
Scale: 10: Full Red, 8: 75 % Red, 6: 50 % Red, 4: 25 % Red, 2: green; whereas, numbers 1, 3, 5, 7, and 9 were used to represent 100 % green, less than 25 %, 50 %, 75 % and 100 % red color of tomato fruit respectively (Wills et al., 2007).
2.8 Statistical analysis
The data was analyzed using Statistix 8.1, a statistical package. Tukey HSD test was performed to evaluate the treatment means at P < 0.05 (n = 5). Origin 2023 software (Origin Lab, Massachusetts, USA) was used for the visualization of data and found the association between different growth attributes, antioxidant enzymatic activities, yield, and quality parameters.
3 Results
3.1 Shoot attributes
Shoot length, shoot fresh biomass, and shoot dry biomass (Table 1) of both heat tolerant (T60 F1 and Super cash F1) and heat sensitive (Naqeeb and Nagina) genotypes of tomato were improved significantly (p ≤ 0.05) by foliar spray of melatonin as compared to control. However, foliar application of melatonin enhanced shoot attributes more in heat tolerant cultivars of tomato as compared to heat sensitive.
Treatments
Tolerant Cultivars
Sensitive Cultivars
T60 F1
Super cash F1
Naqeeb
Nagina
Shoot length (cm)
Control
85.82d
93.41c
65.28f
63.53 g
25 µM
118.98b
131.90a
83.38e
82.34e
Shoot fresh biomass (g)
Control
333.67c
320.32d
244.55e
249.72e
25 µM
468.99a
450.81b
317.23d
331.08
Shoot dry biomass (g)
Control
92.90d
97.35c
52.47 h
57.89 g
25 µM
127.52b
133.03a
65.92f
74.16e
Root length (cm)
Control
47.97d
43.08f
38.49 g
33.27 h
25 µM
66.23a
61.38b
49.82c
43.93c
Root fresh biomass (g)
Control
124.88e
139.47d
113.95f
101.68 g
25 µM
180.44b
197.38a
153.17c
140.26d
Root dry biomass (g)
Control
33.93d
37.12c
21.78 g
21.28 g
25 µM
47.37b
53.13a
28.45e
27.05f
Number of Leaves plant−1
Control
32.51 cd
31.50d
21.96 g
25.52f
25 µM
45.84a
43.69b
27.79e
34.01c
Numbr of Flowers plant−1
Control
40.68c
36.91d
30.99e
27.12f
25 µM
58.38a
54.08b
41.77c
35.76d
3.2 Root attributes
The data regarding root attributes (Table 1) indicated that the foliar spray of melatonin significantly (p ≤ 0.05) improved root fresh biomass, root dry biomass, and root length of both heat tolerant (T60 F1 and Super cash F1) and heat sensitive (Naqeeb and Nagina) genotypes of tomato grown under field conditions. However, heat-tolerant genotypes of tomatoes responded better than heat-sensitive ones.
3.3 Number of leaves and flowers plant−1
Likewise, a significant (p ≤ 0.05) increase in counting of number of leaves plant−1 and number of flowers plant−1 (Table 1) was observed on both heat tolerant (T60 F1 and Super cash F1) and heat sensitive (Naqeeb and Nagina) genotypes of tomato when melatonin spray was applied over untreated control. Overall, the heat-tolerant genotype showed better results as compared to heat-sensitive cultivars.
3.4 Antioxidant enzyme activity
A significant (p ≤ 0.05) increase in the production of enzymes such as SOD, POD, and CAT (Fig. 2) was recorded in all cultivars of tomato either intolerant (T60 F1 and Super cash F1) or in sensitive (Naqeeb and Nagina) as compared to their respective control treatment when melatonin spray was applied. However, a better activity of antioxidant enzymes was recorded in heat-tolerant cultivars when compared with heat-sensitive genotypes.
The percent rise in SOD contents was 47.25 % and 49.71 % in T60 F1 and Super cash F1 respectively whereas 36.81 % and 34.62 % in Naqeeb and Nagina respectively over control treatment. The percentage of increase in POD activity was observed as 50.12 % and 48.47 % in T60 F1 and Super cash F1 respectively and 35.48 % and 37.36 % in Naqeeb and Nagina respectively as compared to control. In the same way, the rise in CAT contents was 37.44 % and 39.11 % in T60 F1 and Super cash F1 respectively, and 25.27 % and 22.75 % in Naqeeb and Nagina respectively when compared with their respective control.
3.5 Osmolytes and total soluble sugars
Results regarding osmolytes (proline and MDA contents) and TSS (Figs. 2 and 3) in all tomato cultivars reflected that foliar spray of melatonin improved proline contents, MDA contents, and total soluble sugars significantly (p ≤ 0.05) as compared to their respective control treatments in both heat tolerant (T60 F1 and Super cash F1) and heat sensitive cultivars (Naqeeb and Nagina) of tomato. The percentage of increase in proline contents was 54.24 % and 50.31 % in T60 F1 and Super cash F1 respectively and 39.17 % and 42.35 % in Naqeeb and Nagina respectively. Similarly, MDA contents were increased by 38.94 % and 35.31 % in T60 F1 and Super cash F1 respectively, and 30.27 % and 27.55 % in Naqeeb and Nagina respectively as compared to control. Similarly, total soluble sugars were increased with the percent values of 35.34 % and 39.74 % in T60 F1 and Super cash F1 respectively, and 26.67 % and 28.90 % in Naqeeb and Nagina respectively over control. However, it was noted that foliar application of melatonin enhanced the Proline contents, MDA, and TSS in heat-tolerant genotypes of tomato as compared to heat-sensitive.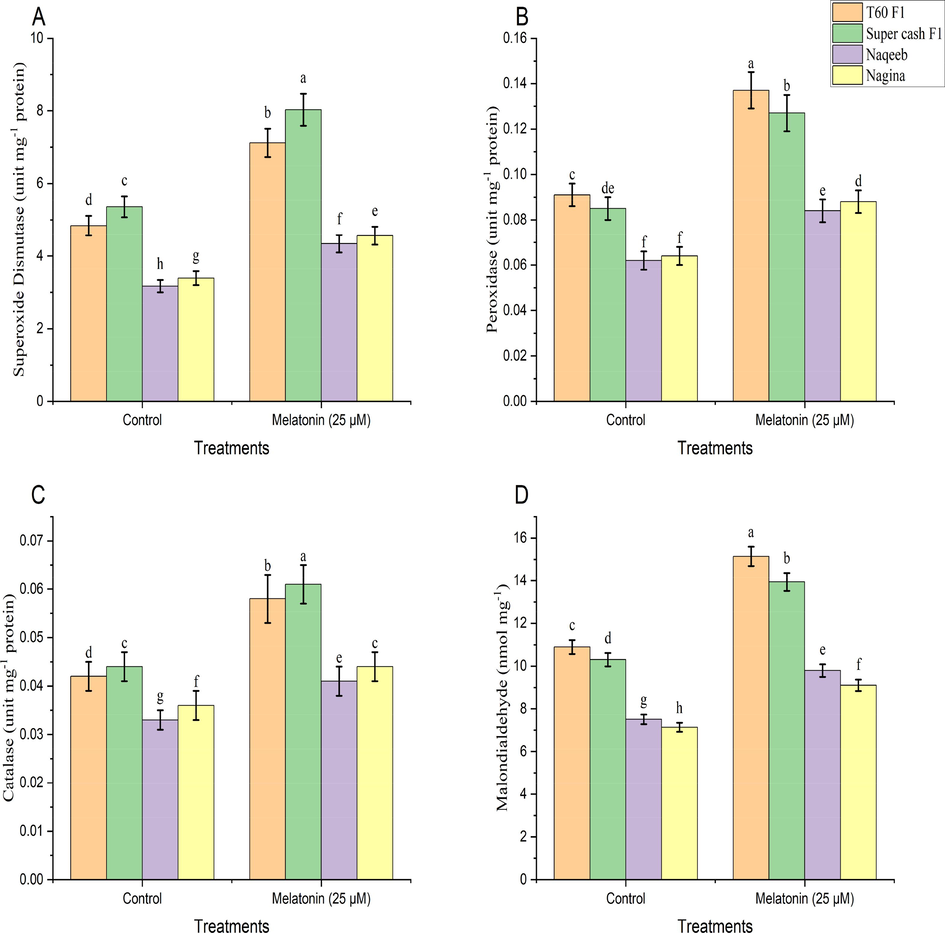
Superoxide dismutase (A), peroxidase (B), catalase (C) and malondialdehyde (D) of different genotypes tomato (Solanum lycopersicum L.) subjected to foliar spray of melatonin under heat stress conditions (Mean ± S.E.).
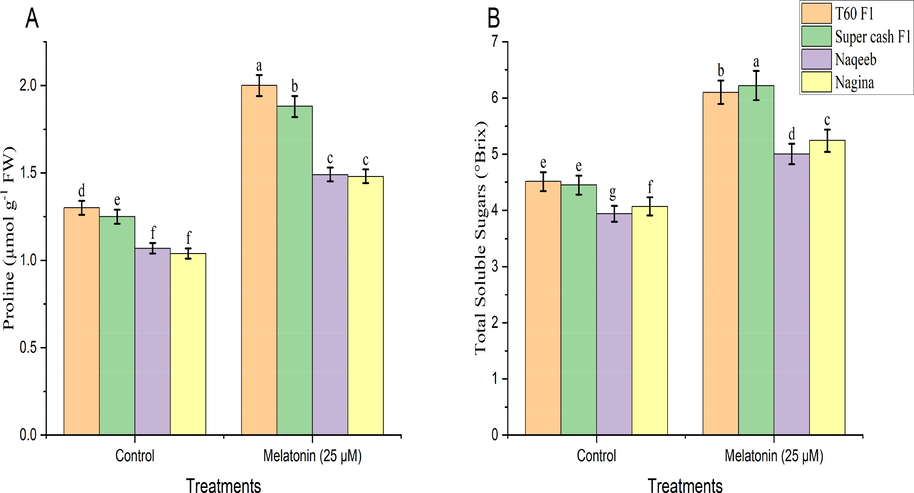
Proline (A) and total soluble sugars (B) concentration of different genotypes tomato (Solanum lycopersicum L.) subjected to foliar spray of melatonin under heat stress conditions (Mean ± S.E.).
3.6 Quality and yield attributes
The results regarding quality and yield attributes (Table 2) in terms of the number of fruits plant -1, fruit size, fruit weight, fruit diameter, fruit color, pH, and yield pot−1 were improved significantly (p ≤ 0.05) owing to foliar spray of melatonin on both heat tolerant (T60 F1 and Super cash F1) and heat sensitive (Naqeeb and Nagina) tomato cultivars as compared to their respective control treatments. However, heat-tolerant genotypes showed more improvement in quality and yield as compared to heat-sensitive cultivars.
Treatments
Tolerant Cultivars
Sensitive Cultivers
T60 F1
Super Cash F1
Naqeeb
Nagina
Number of Fruits plant−1
Control
22.80d
22.88c
13.20 h
15.00 g
25 µM
30.40b
32.40a
17.20f
20.20e
Fruit Size (mm)
Control
76.07f
77.95e
67.62 h
69.50 g
25 µM
99.91b
105.47a
82.75d
86.56c
Fruit weight (g)
Control
101.43d
109.88c
80.77f
87.34e
25 µM
141.34b
150.99a
103.55d
110.68c
Fruit diameter (mm)
Control
60.11d
63.33c
51.94f
53.53e
25 µM
76.00a
81.68a
59.87d
63.16c
Fruit colour
Control
9.33b
9.03b
7.51c
6.57d
25 µM
9.91a
9.88a
7.78c
6.88d
Yield plant−1 (kg)
Control
5.73e
6.39c
4.51 g
4.81f
25 µM
8.33b
9.39a
6.07d
6.51c
pH of Tomato
Control
4.23c
4.41b
3.13 g
3.91f
25 µM
4.41b
4.57a
3.97e
4.05d
3.7 Pearson association between growth parameters, antioxidants activity, osmolytes, TSS, and quality attributes of two different genotypes of tomato
The data revealed that the shoot fresh, shoot dry weight, shoot length, root fresh weight, root dry weight, root length, and dry weight, number of leaves plant-1, number of flowers plant-1, proline contents, MDA contents, quality attributes were significantly and positively associated with POD, SOD, and CAT an in all the tested genotypes of tomato (Fig. 4).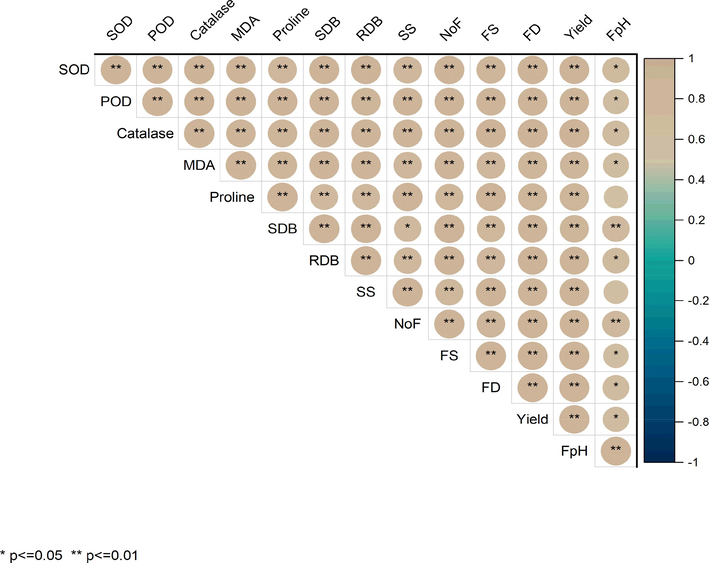
Correlation among growth attributes, antioxidant enzymes activity, proline contents, MDA contents, total soluble sugars (TSS) and quality attributes of different tomato cultivars.
4 Discussion
Heat stress reduced growth attributes in both genotypes of tomatoes and this adverse impact of heat stress was mitigated by foliar application of melatonin. The plants treated with foliar spray of melatonin showed improvement in shoot and root length, shoot and root fresh biomass, shoot and root dry biomass, number of leaves, and number of flowers per plant as compared to control (without melatonin sprayed plants) (Martinez et al., 2018). This might be due to the role of melatonin in increased cell division (Zhang, et al., 2015; Li et al., 2017a) and cell elongation (Kocal et al., 2008) as well as in restricted production of reactive oxygen species (Martinez et al., 2016). Both these factors are linked with an increase in chlorophyll contents and the overall growth of tomatoes. Heat stress also reduced water contents in the leaf tissues causing necrosis, melatonin foliar spray enhanced the water absorbing capacity of the cell by modifying the permeability of the cell membrane and cell wall (Kumar et al., 2014) led to improved membrane stability index and subsequently the various growth attributes. Furthermore, melatonin also regulates the movement and transport of different hormones like gibberellins, auxins, and cytokinins, which are associated with the movement of nutrients and carbohydrates from above-ground portion to underground parts. Thus, it changes the cellular sensitivities of many growth promoters, resulting in the growth of plants getting increased (Martinez et al., 2018).
Heat stress consequences in the production of ROS species which actuate the various phototoxic reactions like protein degradation, lipid peroxidation (LPO), and alteration in DNA (Pospisil and Yamamoto, 2017). Plants sprayed with melatonin presented greater activities of SOD, POD, and CAT enzymes in both heat-tolerant and heat-sensitive genotypes of tomatoes. The increased activity of antioxidant enzymes was due to the role of melatonin in scavenging ROS species and the cellular redox homeostasis mechanism. Hence, melatonin significantly eliminated the drastic impact of heat stress by minimizing the ROS species and lipid peroxidation with the help of antioxidant enzymes (Martinez et al., 2018; Reiter et al., 2007; Kumar et al., 2012; Tan et al., 2015; Wang et al. 2016b; Zhao et al., 2019). Hence, melatonin also augmented antioxidant enzyme activity due to its quality of electron donor to ROS species leading to reduced oxidative damage.
A foliar spray of melatonin significantly raised the accumulation of osmolytes like proline, MDA and total soluble sugars, etc. The rise in production of osmolytes and TSS in this study was an indication of osmotic adjustment owing to increased leaf water contents after the application of melatonin. It indicates that melatonin indirectly accelerates plant growth by reducing the osmotic potential. Other than osmotic adjustments, osmolytes execute different other jobs like the uptake of water, the balance of nutrients, preservation of cell integrity, and regulation of the turgidity of cell (Rivero et al., 2014). Martinez et al. (2016) revealed that melatonin enhances osmolyte contents and soluble sugars in plants grown under abiotic stress, hence responsible for high osmotic adjustment.
Heat stress reduced the quality and yield of tomato plants by inducing ROS species, reducing water contents, and disrupting membrane stability, and photosynthetic apparatus. All these traits are responsible for improving the yield and quality of tomatoes (Hassan et al., 2022). These quality and yield attributes were improved by foliar spray of melatonin because melatonin is considered a master regulator and defensive player in stressful environment and plays its leading role in increasing water contents, protein contents, amino acids, and photosynthetic pigment. All these substances collectively improved the quality and yield of tomatoes in terms of fruit weight, fruit size, fruit diameter, fruit color, pH of tomato, number of fruits plant-1, and yield of fruit pot-1 (Jahan et al., 2019).
5 Conclusion
Heat stress increased the overproduction of ROS species leading to damage the plant defense system. This study demonstrated that the exogenous application of melatonin can mitigate the adverse effects of heat shock by providing a conducive environment for the growth of both types of tomato cultivars under heat stress. Melatonin governs the mechanisms linked with ion homeostasis and improved water absorption, photosynthetic pigment, osmotic adjustment, and nutrient assimilation particularly nitrogen which is the producer of protein and amino acids within the plant. It led to enhanced growth, quality, and defense mechanism by controlling activities of antioxidant enzymes such as SOD, POD, and CAT for the detoxification of ROS species within plants against heat stress. However, the response of heat-tolerant genotypes was promising as compared to heat-sensitive genotypes of tomato plants. Moreover, this research was limited to a pot experiment that provides baseline information about the melatonin-induced heat stress tolerance in tomatoes and will be further evaluated in the field environment of different ecological zones where the elevated temperature is a serious concern.
Declarations
Ethics approval
Not applicable
Consent to participate
All authors consent to participate in the manuscript publication
Consent for publication
All authors approved the manuscript to be published.
Acknowledgement
The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2024R236) King Saud University, Riyadh, Saudi Arabia.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Revisiting the role of ROS and RNS in plants under changing environment. Environ. Exp. Bot.. 2019;161:1-3.
- [Google Scholar]
- Response of tomato (Solanum lycopersicum L.) genotypes to heat stress using morphological and expression study. Plants.. 2022;11(5):615.
- [Google Scholar]
- Melatonin alleviates salt damage in tomato seedling: A root architecture system, photosynthetic capacity, ion homeostasis, and antioxidant enzymes analysis. Sci. Hortic.. 2021;285:110145
- [Google Scholar]
- Melatonin: first-line soldier in tomato under abiotic stress current and future perspective. Plant. Physiol. Biochem.. 2022;185:188-197.
- [Google Scholar]
- AOAC (Association of Official Agricultural Chemistry). (2005). Official Methods of Analysis International, 18th ed.; AOAC: Gaithersburg, MD, USA, 2005.
- Melatonin: a new plant hormone and/or a plant master regulator? Trends. Plant. Sci.. 2019;24(1):38-48.
- [Google Scholar]
- Role of melatonin in alleviating cold stress in Arabidopsis thaliana. J. Pineal. Res.. 2014;56(3):238-245.
- [Google Scholar]
- Rapid determination of free proline for water-stress studies. Plant. and Soil.. 1973;39:205-207.
- [Google Scholar]
- Auxin response factors (ARFs) are potential mediators of auxin action in tomato response to biotic and abiotic stress (Solanum lycopersicum) PLoS One.. 2018;13(2):e0193517.
- [Google Scholar]
- Melatonin mediates enhancement of stress tolerance in plants. Int. J. Mol. Sci.. 2019;20(5):1040.
- [Google Scholar]
- Involvement of antioxidants and lipid peroxidation in the adaptation of two cool-season grasses to localized drought stress. Environ. Exp. Bot.. 2001;45:105-114.
- [Google Scholar]
- Superoxide dismutase I. Occurrence in higher Plants. Plant Physiology.. 1977;59:309-314.
- [Google Scholar]
- Does melatonin improve the yield attributes of field-droughted banana under Egyptian semi-arid conditions? J. Water and Land Development. 2022
- [Google Scholar]
- Melatonin alleviates heat-induced damage of tomato seedlings by balancing redox homeostasis and modulating polyamine and nitric oxide biosynthesis. BMC Plant Biol.. 2019;19(1):1-16.
- [Google Scholar]
- Integrative roles of nitric oxide and hydrogen sulfide in melatonin-induced tolerance of pepper (Capsicum annuum L.) plants to iron deficiency and salt stress alone or in combination. Physiol. Plant.. 2020;168(2):256-277.
- [Google Scholar]
- Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere.. 2019;225:627-638.
- [Google Scholar]
- Cell wall-bound invertase limits sucrose export and is involved in symptom development and inhibition of photosynthesis during compatible interaction between tomato and Xanthomonas campestris pv Vesicatoria. Plant Physiol.. 2008;148:1523-1536.
- [Google Scholar]
- Protection against heat stress in wheat involves change in cell membrane stability, antioxidant enzymes, osmolyte, H2O2 and transcript of heat shock protein. Int. J. Plant Physiol. Biochem.. 2012;4:83-91.
- [Google Scholar]
- 28-homobrassinolide- induced exaggerated growth, biochemical molecular aspects of Brassica juncea L. RLM-619 seedlings under high temperature stress. J. Plant Biochemistry and Physiology.. 2014;2:127-137.
- [CrossRef] [Google Scholar]
- Exogenous melatonin confers salt stress tolerance to watermelon by improving photosynthesis and redox homeostasis. Frontier in Plant Sciences.. 2017;8:295.
- [Google Scholar]
- Accumulation of flavonols over hydroxycinnamic acids favors oxidative damage protection under abiotic stress. Frontier in Plant Sciences.. 2016;7:838.
- [Google Scholar]
- Tolerance to stress combination in tomato plants: new insights in the protective role of melatonin. Molecules.. 2018;23:535.
- [CrossRef] [Google Scholar]
- Glutathione homeostasis as an important and novel factor controlling blossom-end rot development in calcium-deficient tomato fruits. J. Plant Physiol.. 2012;169:1719-1727.
- [Google Scholar]
- Transcriptional regulatory network of plant heat stress response. Trends Plant Sci.. 2017;22(1):53-65.
- [Google Scholar]
- Melatonin and its metabolites: new findings regarding their production and their radical scavenging actions. Acta Biochim. Pol.. 2007;54:1-9.
- [Google Scholar]
- The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environment.. 2014;37:1059-1073.
- [Google Scholar]
- Potential roles of melatonin and sulfur in alleviation of lanthanum toxicity in tomato seedlings. Ecotoxicol. Environ. Saf.. 2019;180:656-667.
- [Google Scholar]
- Melatonin and calcium function synergistically to promote the resilience through ROS metabolism under arsenic-induced stress. J. Hazard. Mater.. 2020;398:122882
- [Google Scholar]
- Melatonin as a potent and inducible endogenous antioxidant: synthesis and metabolism. Molecules.. 2015;20(10):18886-18906.
- [Google Scholar]
- The regulatory effect of melatonin on physiological, biochemical and molecular parameters in cold-stressed wheat seedlings. Plant Growth Regul.. 2014;74:139-152.
- [Google Scholar]
- Exogenous melatonin improves growth and photosynthetic capacity of cucumber under salinity-induced stress. Photosynthetica.. 2016;54:19-27.
- [Google Scholar]
- Post harvest an introduction to thephysiology and handling of fruits and vegetables and ornamentals. University of New South Wales Press. 2007
- [Google Scholar]
- Exogenous melatonin improves salt tolerance in tomato by regulating photosynthetic electron flux and the ascorbate–glutathione cycle. J. Plant Interact.. 2019;14(1):453-463.
- [Google Scholar]
- Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot.. 2015;66:647-656.
- [Google Scholar]
- Effects of melatonin on seedling growth, mineral nutrition, and nitrogen metabolism in cucumber under nitrate stress. J. Pineal Res.. 2017;62(4):e12403.
- [Google Scholar]
- Melatonin synthesis and function: evolutionary history in animals and plants. Frontiers in Endocrinology (lausanne).. 2019;10:249.
- [Google Scholar]
Further Reading
- Melatonin treatment reduces chilling injury in peach fruit through its regulation of membrane fatty acid contents and phenolic metabolism. Food Chem.. 2018;245:659-666.
- [Google Scholar]
- K.A. Gomez A.A. Gomez Statistical Procedures for Agricultural Research, 2nd ed. John 430 Wiley and Sons 1984 New York.
- Improving water-use efficiency by decreasing stomatal conductance and transpiration rate to maintain higher ear photosynthetic rate in drought-resistant wheat. The Crop Journal.. 2017;5(3):231-239.
- [Google Scholar]




