

Translate this page into:
Drought-induced changes in plant-yield interlinked biochemistry of cauliflower (Brassica oleracea L. var. botrytis) by exogenously applied alpha-tocopherol
⁎Corresponding author. drnudrataisha@gcuf.edu.pk (Nudrat Aisha Akram)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
To assess the potential of exogenously-applied vitamin E commonly known as alpha-tocopherol (α-TOC) in drought stress tolerance, an experiment was performed on cauliflower (Brassica oleracea L. var. botrytis) plants. Two cultivars (TCF-605 and White Figer) of cauliflower were subjected to control and drought stress conditions. After thirty days of drought stress treatment, cauliflower plants were treated with α-TOC (150 and 300 mg/L) as a foliage spray. Drought stress markedly inhibited the plant biomass, leaf characteristics such as relative water contents (RWC), chlorophyll pigments (a and b), proline, ascorbic acid (AsA), GB, however it elevated RMP, total phenolics, activities of antioxidant enzymes e.g., CAT, SOD and POD. Drought stress also adversely affected curd characteristics including its fresh weight, GB, proline, total phenolics and enzymes activities (SOD and CAT). Exogenously applied α-TOC considerably suppressed RMP, while it improved plant growth, yield, RWC, total phenolics, ascorbic acid and POD, SOD, and CAT enzyme activities in the leaves of the cauliflower. Exogenous application of α-TOC also raised curd total phenolics, AsA and the activities of all observed enzymes under water deficiency. The cv. TCF-605 was better in shoot weight, shoot length, RWC, proline as well as GB contents, while cv. White Figer was higher in leaf RMP, proline and AsA levels under drought stress conditions. Overall, the results exhibited that cauliflower cv. TCF-605 was relatively tolerant to water stress in relation to growth and yield outcomes, which was interlinked with α-TOC-induced improved leaf and curd osmoprotection and antioxidative defense system. So, exogenous application of α-TOC can be suggested for improving growth and yield potential of crops subjected to drought stress.
Keywords
Drought tolerance
Cauliflower
Osmoprotection
Alpha-tocopherol
Antioxidative defense system

1 Introduction
Water scarcity is a barrier to the global agricultural economy because of the complexity of the water-limited environment and climatic change (Asghar et al., 2022). Drought is defined as a period of low rate of precipitation due to low rainfall or relatively high transpiration rate which results reduction in crop production. It considered multi-facet stress that influence many stages of development including germination, seedling, reproductive and grain/fruit formation. Water flow towards elongating cells is interrupted when there is a severe water shortage, which inhibits cell elongation. It can have enormous effects as its severity and duration progressively rise, making it one of the important adversaries. In dehydration, the turgor pressure and water potential of plants are diminished and plant cells unable to manage their normal metabolism functioning including photosynthesis, respiration, nutrients and water uptake (Badr and Brüggemann, 2020). A decrease in RWC is the preliminary impact of water stress on plants. Stomatal closure is caused by reduced RWC as water pressure/potential decreases under stress. Water stress declines photosynthetic pigments due to reduction of efficiency of PSII, photochemical (QP), and quantum yield (ΦPSII). Drought stress significantly decreases gas exchange attributes (E, A, and Pn) while oxidative damage was observed in tomato plants (Raja et al., 2020).
Brassica oleracea commonly called cauliflower, var. botrytis is a common vegetable that is widely grown across the world. Cauliflower has a delicate flavor and is more versatile for cooking than that white-curd cabbage. Cauliflower fruits are shoots with embryo inflorescences containing several beneficial nutrients. Cauliflower ranks highest among all cabbage varieties in terms of nutrient contents, digestibility, and flavor. Cauliflower has 1.5 to 2.0 times more protein contents in contrast to white-curd cabbage, and 2–3 times amount of ascorbic acid and alkaline mineral salts (Pusik et al., 2018). Cauliflower generally grows best in a cool and moist environment and is susceptible to extremely low temperatures. The optimum growth requires maximum, 15.4–18.9 °C and minimum, 4.6––6.8 °C temperature. Cauliflower plants require excess water during the period when their florets are at their maximum growth before harvest. Due to lack of water, low-quality florets begin to grow early. It is also among the key factors affecting crop yield. The resulting water shortage is usually apparent by decline in intensity of growth, followed by reduction in CO2 assimilation (Hnilička et al., 2010). Cauliflower is sensitive to water stress since it requires 380 to 500 mm of water with irrigation intervals of 5 to 10 days and 45 % rhizosphere depletion permissible throughout growth (Khan et al., 2018).
Exogenous supplementation of antioxidants has recently gained a ground as a possible method for decreasing adversaries of drought on the development and metabolism of plants (Sadiq et al., 2019). Tocopherols are water-insoluble, non-enzymatic antioxidants that belong to family of vitamin E. Green photosynthetic plants naturally produce tocopherols. Tocopherol biosynthesis occurs in different parts of protoplast, chloroplast and vacuole etc. (Li et al., 2008). Only α-tocopherol is found in the membrane of the chloroplast (Grilo et al., 2014), whereas gamma-TOC is mostly present in seeds, nuts, and fruits. Tocopherols are abundantly present in membranes, they prevent membrane lipid peroxidation and are the scavengers of free radicals (Munné‐Bosch, 2007). Under abiotic stresses, tocopherols are crucial for phloem loading and protecting photosystem II against photo-inhibition (Hyun et al., 2011). Plants with insufficient quantity of α-TOC seedlings' growth, impaired photo-assimilates translocation, and accelerated leaf fall (Sattler et al., 2004). Under drought stress, one molecule of alpha-tocopherol is thought to be able to deactivate 120 molecules of oxygen. All other species rely on photosynthetic organisms for their tocopherol production. Tocopherols regulate gene expression in addition to protecting cellular membranes (Munné‐Bosch, 2007). The antioxidant abilities of tocopherols are linked to protons of free radicals (Grebowski et al., 2020). α-TOC is the most dominant antioxidant among the several forms of tocopherols, whilst beta- and gamma-tocopherols have a moderate and delta-TOC with the least antioxidant potential (Kapoor et al., 2015). The hypothesis of the study was whether exogenously applied alpha-tocopherol regulates drought tolerance potential of cauliflower plants and yield production. Thus, the primary objectives of the current experiment were to observe the changes in growth, and physio-biochemical characteristics of cauliflower plants subjected to drought stress. The influence of alpha-tocopherol in mending the osmoprotectant and defense potential against oxidative stress of cauliflower under drought stress was assessed. In addition, the physio-biochemical characteristics of curd of drought-stressed cauliflower treated with α-TOC were determined.
2 Materials and methods
To analyze the effectiveness of foliar-applied alpha-tocopherol (α-TOC) on leaf and curd physio-biochemistry of cauliflower, the seedlings of cauliflower cultivars namely TCF-605 and White Figer were obtained from the AARI, Faisalabad, Pakistan. A total of 36 plastic pots were prepared each with 7.5 kg of soil and five seedlings were transplanted in each pot. The soil used was sandy-loam having pH, 7.8; EC, 2.1 dS/m; organic matter, 3.1 % and saturation percentage was 39 %. The experiment was planned in CRD with replicates of three each of drought stress or α-TOC. During the experiment, relative humidity was 59 ± 2; sunshine, 7.5 h and average day temperature 26 ± 1.4 °C. The seedlings were established properly before the initiation of drought stress treatments. The drought stress (60 % F.C.) along with control (100 % F.C.) were maintained 20 days after transplanting the 42-day-old seedlings. The drought stress treatment was maintained for 30 days. After this specific period, varying concentrations of α-TOC (150 and 300 mg/L) were applied as foliage application by using manual plastic sprayer. After two weeks of α-TOC supplement, plants were used for sampling fresh leaves and preserved in a freezer at −20 °C. Two plants were harvested carefully along with roots. Then they were detached into roots and shoots, determined their lengths and weights (fresh and dry). After sun-drying, these samples were dried at 60 °C using an oven for three days and noted dry weights. Remaining plants were allowed to grow till curd formation for the determination of yield as well as different physio-biochemical attributes of cauliflower:
2.1 Relative water contents (RWC)
A fresh third leaf from top was harvested and noted their fresh weight. Then, dipped the leaves into water for 3 h and noted their turgid weight. After removing from water, air dried samples were oven dried at 70 °C till their constant dry weight were obtained. The RWC worked out following Jones and Turner (1978).
2.2 Relative membrane permeability (RMP)
A fresh third leaf (0.5 g) from the top of a plant was detached and chopped in deionized water (10 mL). After noting the initial electrical conductivity (EC°), the samples were put at 4 °C for night and their EC1 was noted. After that, the leaves were heated for 15 min, and cooled at room temp and noted EC2 (Yang et al., 1996).
2.3 Chlorophyll pigments
Following the procedure proposed by Arnon (1949), fresh leaf and curd samples (each 0.25 g) were extracted in 5 mL of acetone (80 %; v/v).
2.4 Free proline contents
Fresh leaf or curd (0.25 g) was ground in 5 mL sulfosalicylic acid (3 %) and filtered. The homogenate was mixed with acid ninhydrin and boiled the mixture at 100 °C for one hour. After boiling, mixture was cooled in an ice bath. Reaction mixture was mixed with 5 mL of toluene in each test tube. After vortexing for one minute, two clearly distinct layers were formed. The absorbance of upper layer was recorded at 520 nm following Bates et al. (1973).
2.5 Glycinebetaine (GB)
Fresh samples (0.25 g) were extracted each with 5 mL d·H2O, and centrifuged at 10,000 x g for 10 min. After that, 1 mL supernatant from upper layer taken and mixed with 1 mL H2SO4 and 0.2 mL of KI3. Cooled for 90 min and added 2.8 mL distilled water to 6 mL of 1, 2 dichloroethane, resulted in the formation of two layers. Then separated the lower layer and absorbance was recorded at 365 nm followed by Grieve and Grattan (1983).
2.6 Total phenolics
A sample (0.1 g) of fresh leaf or curd was homogenized with 5 mL 80 % acetone and centrifuged at 10, 000 x g for 10 min. The final reading was noted at 750 nm (Julkunen-Tiitto, 1985).
2.7 Ascorbic acid
Fresh leaf and curd samples (each 0.25 g) were extracted using TCA (10 mL; 6 %). Then, 2 mL of 2 % DNPH was mixed in 4 mL sample extract, and 1 mL thiourea was mixed followed by Mukherjee and Choudhuri (1983).
2.8 Antioxidant enzymes
Fresh samples (each 0.25 g) were extracted in ice-cooled 5 mL buffer (KH2PO4; pH 7.8) and preserved the samples in Eppendorf tubes. After centrifugation at 10000 x g, assessed the activities of the following antioxidant enzymes:
The POD enzyme activity was determined by using 50 µL enzyme extract, 100 µL H2O2, 40 mM KH2PO4 (50 µL), and guiacol (100 µL) and mixed precisely and OD was determined at 470 nm (Chance and Maehly, 1955).
To assess the activity of SOD enzyme, the reaction mixture was used to observe the absorbance of each sample at 560 nm (Giannopolitis and Ries, 1977).
For CAT activity, leaf and curd extracts (each 0.1 mL) were homogenized with 1 mL of H2O2, and 1.9 mL K buffer and noted OD at 240 nm (Chance and Maehly, 1955).
2.9 Statistical analysis
ANOVA and LSD at 5 % probability level were determined using statistical software, Cohort, Monterey, CA, USA.
3 Results
3.1 Growth attributes
Fresh and dry weights (shoot and root) of both cauliflower cultivars (TCF-605 and White Figer) decreased significantly under drought stress [60 % field capacity (F.C.)] conditions (Fig. 1). Application of α-TOC at the concentration of 150 mg/L and 300 mg/L markedly improved the shoot and root fresh and dry biomass under both water regimes of cauliflower plants. Of varying levels of α-TOC, 300 mg/L was most profound in accelerating all growth attributes. Root fresh and dry weights were same in both cultivars, while cv. TCF-605 was better in shoot (dry and fresh) weights under both watering levels.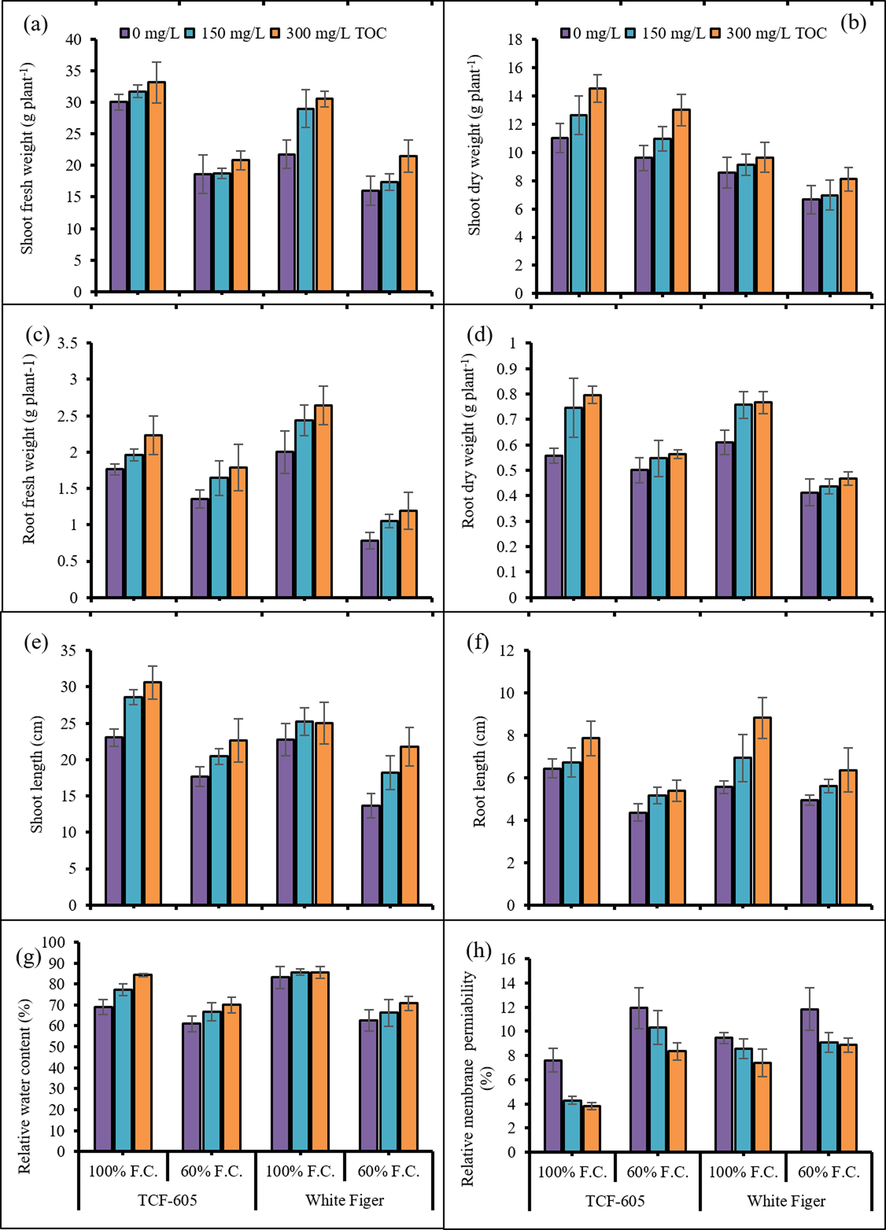
Shoot fresh weight (a), shoot dry weight (b), root fresh weight (c), root dry weight (d), shoot length (e), root length (f), leaf relative water content (g) and leaf relative membrane permeability (h) of two cultivars of cauliflower (Brassica oleracea L. var. Botrytis) subjected to foliar applied alpha-tocopherol under drought stress conditions (Mean ± S.E.).
A considerable suppression was noted in root and shoot lengths of drought-stressed cauliflower plants. However, TOC considerably enhanced (P ≤ 0.01) the shoot and root lengths of cauliflower plants. The cv. TCF-605 was superb in shoot length particularly under control conditions, while root length was persistent in both cauliflower cultivars under varying water levels (Fig. 1).
3.2 Leaf characteristics
Data for relative water contents (RWC) exhibited a considerable decrease in this attribute under drought stress. While α-TOC had a positive influence on RWC particularly in cv. TCF-605 under control conditions (Fig. 1). Overall, the RWC were same in both cauliflower cultivars.
Water stress significantly enhanced the RMP of all observed plants. The foliar-applied α-TOC effectively reduced the RMP under both stress and no stress regimes. The cv. White Figer was relatively higher in RMP under both watering levels.
A notable reduction (Fig. 2) in leaf chlorophyll a and b contents, while an increase in chlorophyll a/b ratio was observed in both cauliflower cultivars under varying water regimes. Exogenously applied α-TOC had no impact on chlorophyll a concentration, while a significant increase was observed in chlorophyll b and ratio of a/b of all cauliflower plants.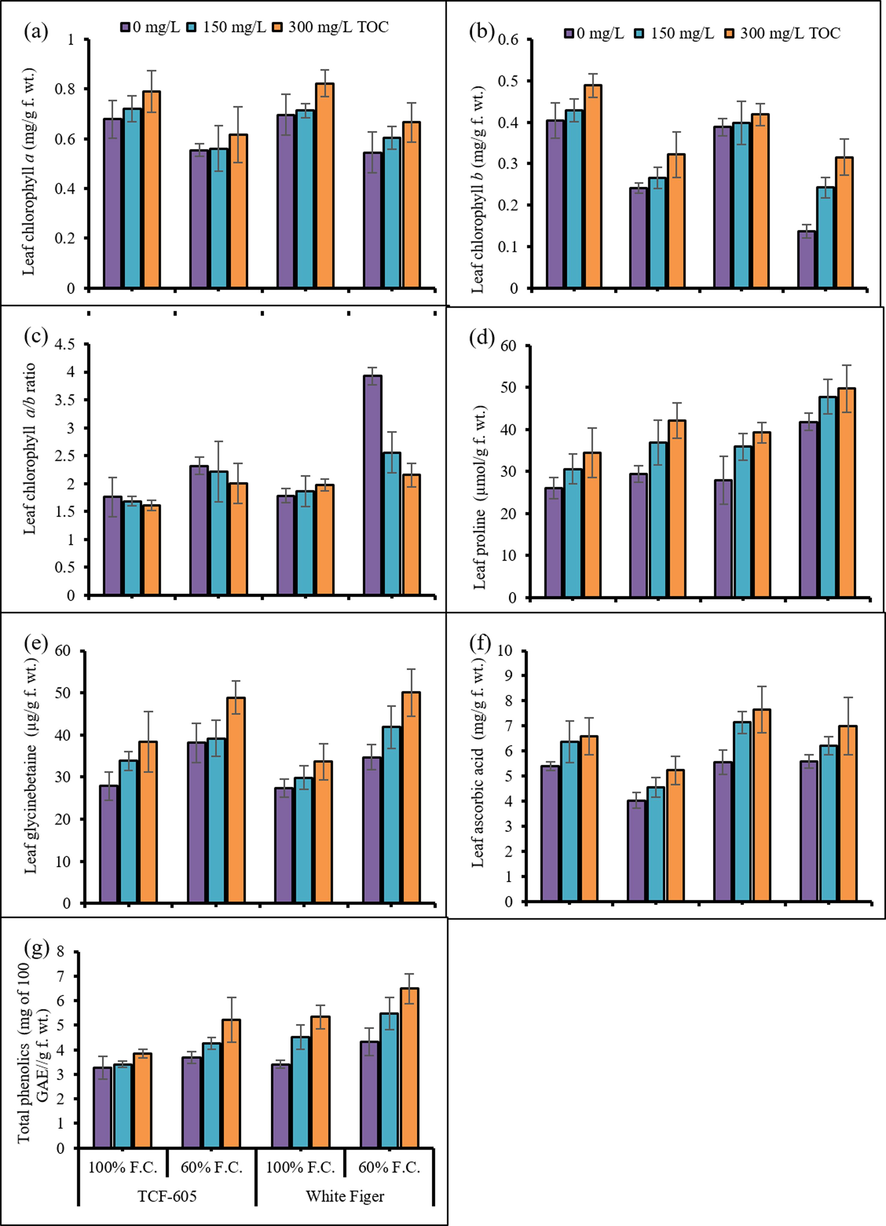
Leaf chlorophyll a (a), chlorophyll b (b), total chlorophyll (c), proline (d), glycinebetaine (e), ascorbic acid (f) and total phenolics (g) of two cultivars of cauliflower (Brassica oleracea L. var. Botrytis) subjected to foliar applied alpha-tocopherol under drought stress conditions (Mean ± S.E.).
A considerable rise was noted in proline and GB concentrations of all cauliflower plants under stress conditions (Fig. 2). Foliar-applied α-TOC markedly enhanced the accumulation of proline as well as GB contents at 60 % F.C. The cv. White Figer was better in accumulating more proline contents than cv. TCF-605. However, leaf GB contents remained almost same in all cauliflower plants under stress conditions.
A marked decrease (P ≤ 0.01) was noticed (Fig. 2) in leaf AsA contents of all cauliflower plants under water scarce regime. Exogenous spray of α-TOC had a considerable improving influence on the ascorbic acid contents of both cauliflower cultivars especially at 60 % F.C. The cv. TCF-605 was lower in AsA contents than the other one under drought stress.
Drought stress effectively (P ≤ 0.01) boosted the total phenolic contents of both cultivars of cauliflower. The treatment of leaves with α-TOC appreciably improved the total phenolic contents particularly at the concentration of 300 mg/L. The cv. White Figer was observed sensitive cultivar in this biochemical attribute under stress regime (Fig. 2).
Activities of antioxidant enzymes including POD (peroxidase), SOD (superoxide dismutase) and CAT (catalase) of all cauliflower plants were found to be significantly accelerated (P ≤ 0.001) under water limited regime. Foliar-applied α-TOC induced a considerable increase in the activities of peroxidase, SOD and CAT enzymes under all water regimes. The cv. TCF-605 showed higher peroxidase activity than that in cv. White Figer. Nonetheless, both cultivars were similar in the CAT and SOD enzymes activities under varying water regimes (Fig. 3).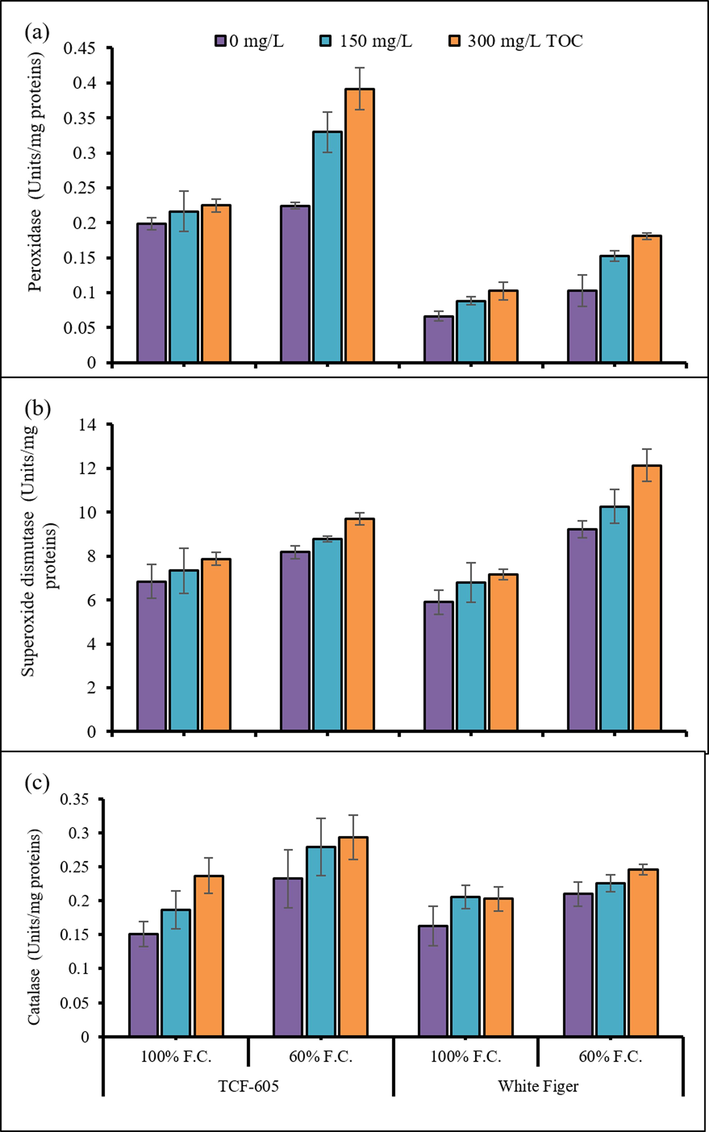
Activities of leaf peroxidase (a), superoxide dismutase (b) and catalase enzymes (c) of two cultivars of cauliflower (Brassica oleracea L. var. Botrytis) subjected to foliar applied alpha-tocopheroler drought stress conditions (Mean ± S.E.).
3.3 Yield characteristics
Water shortage drastically reduced the curd fresh weight of all cauliflower plants. Exogenously applied α-TOC considerably improved (P ≤ 0.05) the curd fresh weight under stress conditions (60 % F.C.). Of both cultivars, cv. TCF-605 showed better results than those of cv. White Figer under varying exogenous treatments and water regimes (Fig. 4).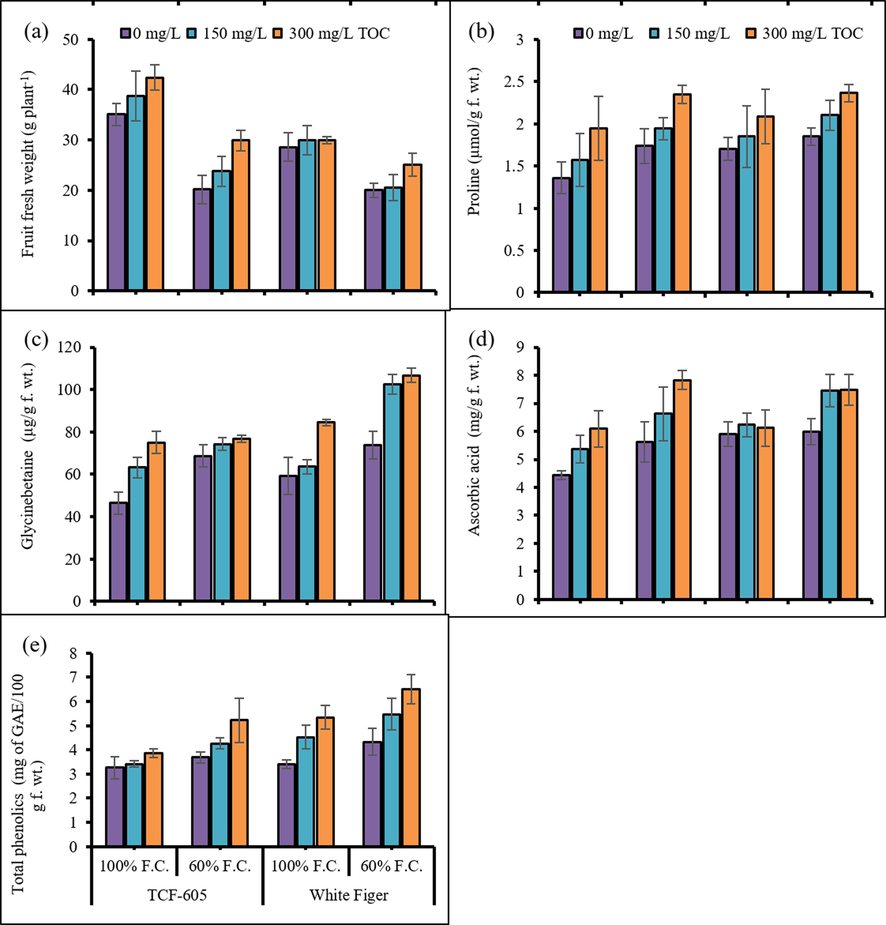
Head fresh weight (a), proline (b), glycinebetaine (c), ascorbic acid (d) and total phenolics (e) of two cultivars of cauliflower (Brassica oleracea L. var. Botrytis) subjected to foliar applied alpha-tocopherol under drought stress conditions (Mean ± S.E.).
The imposition of limited water stress considerably increased the concentrations of curd GB and proline in cauliflower plants. Exogenously applied α-TOC (300 mg/L) indicated effectiveness in terms of enhancing the proline and GB contents in curd tissues of cauliflower cultivars under both water regimes (Fig. 4). The cv. White Figer was relatively higher in curd GB contents than the other cultivar. The curd proline contents were similar in all curds of cauliflower.
Water stress notably raised AsA contents in the curd of both cultivars (Fig. 4). Externally applied α-TOC significantly enhanced (P ≤ 0.01) the curd AsA contents of both cultivars under drought stress conditions. No significant interaction was noted between the cultivars, treatments and drought stress regimes.
A considerable improvement was noted in total phenolic contents in the curds of both cauliflower cultivars under water deficit stress (Fig. 4). Foliar-applied α-TOC had a positive influence on total phenolic contents of all cauliflower plants. Cv. White Figer was better than cv. TCF-605 in total phenolic contents under varying water regimes.
Imposition of water stress markedly increased the activities of POD enzyme, while a significant reduction was noted in SOD and CAT enzymes activities in the curd of both cultivars. Application of α-TOC considerably enhanced (P ≤ 0.05) the activities of POD, SOD and CAT enzymes of both cultivars under varying water regimes. The SOD and POD enzymes activities were similar in the curd of both cauliflower cultivars. However, the CAT activity better in curd of cv. TCF-605 than the other one under both water regimes (Fig. 5).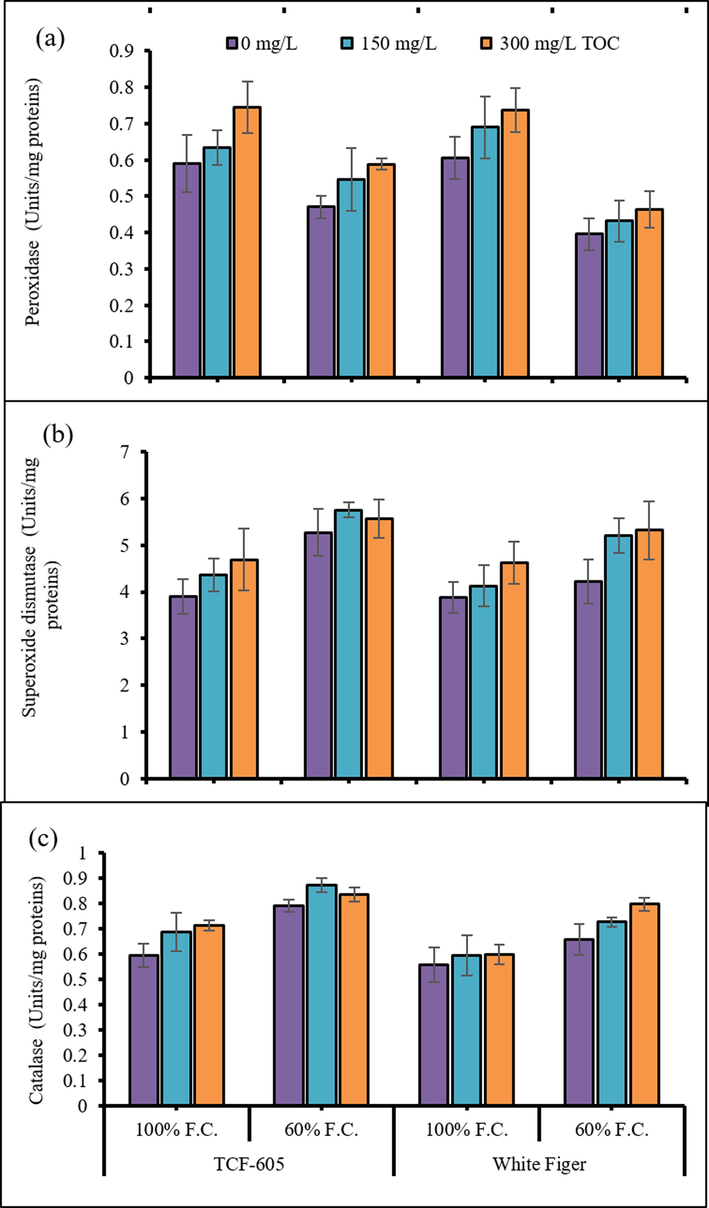
Head activities of peroxidase (a), superoxide dismutase (b), and catalase (c) enzyme of two cultivars of cauliflower (Brassica oleracea L. var. Botrytis) subjected to foliar applied alpha-tocopherol under drought stress conditions (Mean ± S.E.).
4 Discussion
Drought stress alters plant responses including morphological, physiological, and biochemical, which ultimately result in disruption of plant normal development and physiological systems (Kosar et al., 2021). Moreover, water deficiency observed at any growth stage retarded plant growth and yield outcomes, and such growth reduction can be associated with drought-induced osmotic stress, disturbance in photosynthetic rate and efficiency, nutritional and hormonal imbalance and disturbance in protein synthesis. Water stress also generated oxidative stress due to overproduction of ROS, which damage the vital membranes, and trigger acute metabolic disorders, eventually hampering plant yield (Sharma and Zheng, 2019). In this experiment, drought stress (60 % F.C.) inhibited growth attributes of all cauliflower plants. However, externally applied regulator was found best in enhancing plant growth as observed in earlier published studies. For example, Hameed et al. (2021) observed an effective role of foliar-applied α-tocopherol (α-TOC) on plant growth of carrot, and Sadiq et al. (2019) on mung bean plants exposed to water stress regimes. Likewise, exogenous application of α-TOC (150 and 300 mg/L) markedly enhanced all the observed growth characteristics of all cauliflower plants exposed to varying water regimes.
A notable decrease was observed in pigments (chlorophyll a and b) in drought stressed cauliflower plants, According to Ashraf and Harris (2013) stress-induced alterations in chlorophyll biosynthesis or its antagonists may be due to decrease in chlorophyll contents, particularly chlorophyll b contents in both cultivars of cauliflower. However, as described in numerous previous researches (Sadiq et al., 2019), the drought-induced decrease in chlorophyll contents is a well-known adversary. Generally, stress conditions suppressed electron transport rate, efficiency of PSII, alters D1 protein and PPFD which may imbalance electrons, thereby decreased electron transport rate (Guidi et al., 2019). So, appropriate watering was suggested to sustain photosynthetic efficiency and alleviate photo-damage as already recommended for water stressed lily (Zhang et al., 2011), maize (Asghar et al., 2022) and mung bean (Sadiq et al., 2019) plants. However, plants exposed to water stress had higher chlorophyll concentrations after receiving foliar-applied α-TOC. This might have occurred as a result of crucial function of α-TOC in removing lipid peroxyl radicals before they attack to alpha-tocopheroxyl radicals. Approximately a molecule of α-TOC can deteriorate 220 1O2 (Shah et al., 2021).
Water-limited conditions minimized the uptake of water as well as essential nutrients in plant cells/tissues leading to reduced plant yield production. In this study, foliar applied α-TOC considerably enhanced the RWC as well as reduced relative membrane permeability (RMP) under stress conditions. The α-TOC-induced RMP might have prevented cell damage caused by water stress. Under drought stress, membrane lipid-proteins linked with membrane fluidity as well stability. Dynamic changes in leaf membrane composition were examined in wheat (Wang et al., 2020) and carrot (Hameed et al., 2021) under different environmental stresses. The reduced leaf relative water content and electrolyte leakage from cells correspond with the intensity of membranes lipid peroxidation as usually determined by MDA contents. Generally, the lipid peroxidation in terms of MDA contents increased in drought stressed plants irrespective of growth stage. The overproduction of ROS have been involved in lipid peroxidation, photo-oxidative stress, deteriorates vital membranes and proteins as reported earlier in maize (Shafiq et al., 2021).
Osmotic adjustment either by ion homeostasis, osmoregulation, or ionic compartmentalization is a well-known phenomenon involved in water stress tolerance of different crops e.g., canola (Akram et al., 2018), sunflower (Kosar et al., 2021), and maize (Asghar et al., 2022), etc. High proline and GB contents under severe conditions is a key indicator of osmoregulation, which plays a significant role in osmoprotection of plants to counteract stressful conditions (Saadia et al., 2013). Recent reports have suggested the role of proline and GB in scavenging ROS and maintaining water level under stress levels (Asghar et al., 2022). In this study, water shortage and foliar-applied α-TOC significantly enhanced accumulation of proline and GB contents in both leaves and curds of cauliflower plants. Similar findings depicted in rapeseed (B. napus L.) by Bhuiyan et al. (2019) under water stress conditions. In this investigation, high proline and GB suggested to be involved in the oxidative defense system as previously reported that high GB, proline and trehalose upregulated the oxidant defense mechanisms by reducing synthesis of MDA, H2O2 and methylglyoxal contents of drought stressed rapeseed plants. Similarly, in an earlier study with pea (P. sativum L.) plants, Osman (2015) reported high accumulation of GB and proline and both metabolites imparted a significant role in enhancing the activities of enzymatic antioxidants e.g., SOD, APX and CAT under water stress regimes.
In this trial, foliage spray of α-TOC effectively boosted the activities of CAT, SOD, and POD enzymes of cauliflower plants. Antioxidants provide defense against oxidative damage caused by overproduction of ROS. Our examinations corresponded with the suggestions of Sadiq et al. (2017) observed in mung bean. By applying foliar α-TOC during water stress, they found an increase in the enzymatic antioxidants in mung bean plants. The activities of SOD, CAT and POD enzymes were also significantly increased in the curds of cauliflower plants parallel to the findings of Naqve et al. (2021). They found that the antioxidant enzymes in the stressed okra fruits increased significantly. The present study showed that AsA and total phenolics of cauliflower plants enhanced under water stress as well as by exogenously applied α-TOC. So these findings are in accordance to the previous data observed in Achillea species (Gharibi et al., 2016) and sugarcane (Shaoyu et al., 1994).
Water deficiency reduced the yield of cauliflower in terms of curd fresh weight. Water stress-induced reduction in yield production has also been observed in sunflower (Kosar et al., 2021) and maize (Shafiq et al., 2021). External application of α-TOC considerably enhanced the curd fresh weight and overall yield of cauliflower plants which may have been due to increased quantity of photosynthetic pigments, better nutrient uptake, and increased tolerance against oxidative stress. In this study, the exogenously applied α-TOC minimized the damaging effects of drought stress in cauliflower plants.
Overall, drought stress reduced plant growth, RWC, chlorophyll contents, proline, GB, and AsA while increased leaf RMP, total phenolics, and the activities of studied enzymes like peroxidase, SOD, and CAT. It also increased curd fresh weight, proline and GB contents, total phenolics, and antioxidant enzyme activities. While improving plant growth, yield, RWC, total phenolics, AsA, and the activities of enzymatic antioxidants in the leaves of both cauliflower cultivars, exogenously applied α-TOC significantly reduced leaf RMP. Under water stress regimes, α-TOC also enhanced the curd ascorbic acid, activities of all the three enzymes and total phenolics. The cv. TCF-605 was superior than the other one in performance under water stress regime. This may be due to α-TOC-induced upregulation of leaf and curd osmoprotection and antioxidative defense system.
5 Ethics approval
Not applicable.
6 Consent to participate
All authors consent to participate in the manuscript publication
7 Consent for publication
All authors approved the manuscript to be published.
CRediT authorship contribution statement
Nimra Zafar: Methodology. Nudrat Aisha Akram: Conceptualization. Kaneez Fatima: Formal analysis. Sibgha Noreen: Software. Muhammad Sohail Akram: Software, Validation. Saliha Umer: . Muhammad Ashraf: Conceptualization, Data curation. Abdulaziz Abdullah Alsahli: Funding acquisition, Software, Writing – review & editing. Sheikh Mansoor: Funding acquisition, Formal analysis, Validation, Writing – review & editing.
Acknowledgement
The authors would like to extend their sincere appreciation to the Researchers Supporting Project Number (RSP2023R236) King Saud University, Riyadh, Saudi Arabia.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Aminolevulinic acid and nitric oxide regulate oxidative defense and secondary metabolisms in canola (Brassica napus L.) under drought stress. Protoplasma. 2018;255:163-174.
- [Google Scholar]
- Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta Vulgaris. Plant Physiology.. 1949;24(1):1.
- [Google Scholar]
- Shade and microbes enhance drought stress tolerance in plants by inducing phytohormones at molecular levels: a review. J. Plant Ecol.. 2022;15(6):1107-1117.
- [Google Scholar]
- Photosynthesis under stressful environments: an overview. Photosynthetica. 2013;51:163-190.
- [Google Scholar]
- Comparative analysis of drought stress response of maize genotypes using chlorophyll fluorescence measurements and leaf relative water content. Photosynthetica. 2020;58:38-645.
- [Google Scholar]
- Rapid determination of free proline for water-stress studies. Plant and Soil. 1973;39:205-207.
- [Google Scholar]
- Mitigation of PEG-induced drought stress in rapeseed (Brassica rapa L.) by exogenous application of osmolytes. Biocatalysis and Agricultural. Biotechnology. 2019;20:101197
- [Google Scholar]
- Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol.. 2016;178:796-809.
- [Google Scholar]
- Superoxide dismutases: I. Occurrence in Higher Plants. Plant Physiol.. 1977;59(2):309-314.
- [Google Scholar]
- Antioxidant activity of highly hydroxylated fullerene C60 and its interactions with the analogue of α-tocopherol. Free Radic. Biol. Med.. 2020;160:734-744.
- [Google Scholar]
- Rapid assay for determination of water soluble quaternary ammonium compounds. Plant and Soil. 1983;70:303-307.
- [Google Scholar]
- Alpha-tocopherol and gamma-tocopherol concentration in vegetable oils. Food Sci. Technol. (. 2014;34:379-385.
- [Google Scholar]
- Chlorophyll fluorescence, photoinhibition and abiotic stress: does it make any difference the fact to be a C3 or C4 species? Front. Plant Sci.. 2019;10:174.
- [Google Scholar]
- Seed treatment with α-tocopherol regulates growth and key physio-biochemical attributes in carrot (Daucus carota L.) plants under water limited regimes. Agronomy. 2021;11(3):469.
- [Google Scholar]
- Effect of water deficit and application of 24-epibrassinolide on gas exchange in cauliflower plants. Sci. Agric. Bohem.. 2010;41(1):15-20.
- [Google Scholar]
- Role of α-tocopherol in cellular signaling: α-tocopherol inhibits stress-induced mitogen-activated protein kinase activation. Plant Biotechnology Reports.. 2011;5:19-25.
- [Google Scholar]
- Osmotic adjustment in leaves of sorghum in response to water deficits. Plant Physiol.. 1978;61(1):122-126.
- [Google Scholar]
- Phenolic constituents in the leaves of northern willows: methods for the analysis of certain phenolics. J. Agric. Food Chem.. 1985;33(2):213-217.
- [Google Scholar]
- Redox homeostasis in plants under abiotic stress: role of electron carriers, energy metabolism mediators and proteinaceous thiols. Front. Environ. Sci.. 2015;3:13.
- [Google Scholar]
- Impact of exogenously applied trehalose on leaf biochemistry, achene yield and oil composition of sunflower under drought stress. Physiol. Plant.. 2021;172(2):317-333.
- [Google Scholar]
- Current opinions on the functions of tocopherol based on the genetic manipulation of tocopherol biosynthesis in plants. J. Integr. Plant Biol.. 2008;50(9):1057-1069.
- [Google Scholar]
- Implications of water stress-induced changes in the levels of endogenous ascorbic acid and hydrogen peroxide in Vigna seedlings. Physiol. Plant.. 1983;58(2):166-170.
- [Google Scholar]
- Foliar spray of alpha-tocopherol modulates antioxidant potential of okra fruit under salt stress. Plants.. 2021;10(7):1382.
- [Google Scholar]
- Enhancing antioxidant–yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci.. 2015;60(2):389-402.
- [Google Scholar]
- Study into formation of nutritional value of cauliflower depending on the agri-biological factors. Eastern-Eur. J. Enterprise Technol.. 2018;6(11):29-35.
- [Google Scholar]
- Impact of Drought and Heat Stress Individually and in Combination on Physio-Biochemical Parameters, Antioxidant Responses, and Gene Expression in Solanum Lycopersicum. 3 Biotech.. 2020;10:1-18.
- [Google Scholar]
- Comparative study of SOS2 and a novel PMP3-1 gene expression in two sunflower (Helianthus annuus L.) lines differing in salt tolerance. Appl. Biochem. Biotechnol.. 2013;170:980-987.
- [Google Scholar]
- Foliar applications of alpha-tocopherol improves the composition of fresh pods of Vigna radiata subjected to water deficiency. Turk. J. Bot.. 2017;41(3):244-252.
- [Google Scholar]
- Alpha-tocopherol-induced regulation of growth and metabolism in plants under nonstress and stress conditions. J. Plant Growth Regul.. 2019;38:1325-1340.
- [Google Scholar]
- Vitamin E is essential for seed longevity and for preventing lipid peroxidation during germination. Plant Cell. 2004;16(6):1419-1432.
- [Google Scholar]
- Influence of glycine betaine (natural and synthetic) on growth, metabolism and yield production of drought-stressed maize (Zea mays L.) plants. Plants.. 2021;10(11):2540.
- [Google Scholar]
- Effect of exogenous alpha-tocopherol on physiobiochemical attributes and agronomic performance of lentil (Lens culinaris Medik.) under drought stress. PLoS One. 2021;16(8):e0248200.
- [Google Scholar]
- Protective effect of free radical scavengers and drought resistance in sugarcane. Zuo Wu Xue Bao.. 1994;20(2):149-155.
- [Google Scholar]
- Melatonin mediated regulation of drought stress: Physiological and molecular aspects. Plants.. 2019;8(7):190.
- [Google Scholar]
- Dynamic changes in membrane lipid composition of leaves of winter wheat seedlings in response to PEG-induced water stress. BMC Plant Biol.. 2020;20(1):1-15.
- [Google Scholar]
- Effects of high temperature on membrane stability and chlorophyll fluorescence in glycinebetaine-deficient and glycinebetaine-containing maize lines. Funct. Plant Biol.. 1996;23(4):437-443.
- [Google Scholar]
- Effect of water stress on leaf photosynthesis, chlorophyll content, and growth of oriental lily. Russ. J. Plant Physiol.. 2011;58:844-850.
- [Google Scholar]
Appendix A
Supplementary material
Supplementary data to this article can be found online at https://doi.org/10.1016/j.jksus.2023.103028.
Appendix A
Supplementary material
The following are the Supplementary data to this article:




