

Translate this page into:
Changes in biochemical, histological and specific immune parameters in Catla catla (Ham.) by Cynodon dactylon (L.)
*Corresponding author. Mobile: +91 94437 24611; fax: +91 4362265140 drravinikesh@yahoo.co.in (S. Ravikumar)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Available online 31 October 2010
Abstract
Indian major carp, Catla catla (Ham.), was fed with ethanolic extract of Cynodon dactylon (L.) mixed diet with 0.05% (group A), 0.5% (group B) and 5% (group C) extract for 60 days with an interval of 10 days. Significant (P > 0.05) alterations were observed in the haematological and biochemical parameters such as white blood cell counts, red blood cell counts, haemoglobin content, serum glucose, cholesterol, protein, albumin, globulin, albumin/globulin ratio and RNA/DNA ratio in the experimental diet and control fishes. Non-specific immune response was assessed by the anti-protease activity, which was significantly increased in the experimental diet groups. The specific immune response of fish was evaluated by antibody response against heat-killed Aeromonas hydrophila for 28 days by using ELISA and bacterial agglutination assay. Aggregation of melanomacrophage centres (MMC) and considerable modifications were observed in the histological analysis of the spleen of experimental diet groups. These results suggested that C. dactylon (L.) could combat the microbial infection by stimulating the immune response in fish.
Keywords
Cynodon dactylon
Catla catla
Aeromonas hydrophila
Anti-protease
ELISA
Melanomacrophage centres

1 Introduction
Infectious diseases are a major problem in aquaculture causing heavy loss to the fish farmers. The recent expansion of intensive aquaculture practices has led to high interest in understanding the various fish diseases, so that they can be treated or prevented. It is widely demonstrated that the occurrence of diseases in fish farm is due to several factors concerned with the rearing methods, environmental conditions and variations. Consequently, cultivated fish can become more susceptible not only to pathogenic but also to opportunistic bacteria (Woo and Bruno, 1998).
Aeromonas hydrophila is a ubiquitous organism present in the aquatic environment causing diseases in fish under stress (Doukas et al., 1998). A. hydrophila is a gram negative, motile rod recorded as an opportunistic pathogen in freshwater fish species and it is considered to have widespread geographical distribution. The bacterium causes haemorrhagic septicemia, which is characterized by the presence of small superficial lesions, focal haemorrhages particularly in the gills and opercula, ulcers, abscesses, exophthalmia and abdominal distension (Austin and Austin, 1987). It was also reported by Rajeswari et al. (2005) that the isolation of A. hydrophila from dropsy-infected common carp Catla catla and Cirrhinus mrigala has shown the symptoms of distended abdomen, loose scales, deep ulcers on the dorsal surface and extensive haemorrhages on the ventral part of the fishes. Diseases in intensive freshwater aquaculture are closely monitored in India due to economic loss observed in recent years. Recent advancement in immuno-nutrition studies revealed that some nutrients are linked to the immunological status of fish (Priya et al., 2004; Kumar et al., 2005). This has drawn the attention of fish nutritionists to the immunoprotection of fish besides the growth. Sustainable aquaculture depends on perfect balance between growth and health condition of fish. The use of antibiotics and chemotherapeutics to combat fish diseases has the risk of generating resistant pathogens, bioaccumulation and environmental pollution. Commercial vaccines are expensive for fish farming practices and are specific against particular pathogens (Raa et al., 1992).
One of the most promising methods of controlling diseases in aquaculture is strengthening the defense mechanisms of fish through prophylactic administration of immunostimulants (Robertsen, 1999). Several plant materials/products such as Eclipta alba (Christybapita et al., 2007), Aloe vera (Kim et al., 1999), Ocimum sanctum (Logambal et al., 2000), Viscum album, Urtica dioica and Zingiber officinale (Dugenci et al., 2003), Solanum trilobatum (Divyagnaneswari et al., 2007), Astragalus radix and Scutellari radix (Yin et al., 2006), and Achyranthes aspera (Vasudeva and Chakrabarti, 2005a,b) have been reported to enhance the immunity of fish.
Cynodon dactylon (L.) Pers. (Gramineae, Poaceae) is a herbal plant commonly known as ‘Arugampul’ in Tamil Nadu, India, which is treated as a blessing plant. This grass is widely distributed in India and in almost all parts of the world. Traditionally, the juice of this plant is commonly consumed as a health drink during early morning in south India. It takes an important role in Ayurvedic medicine. C. dactylon extract is used to treat hysteria, epilepsy and insanity (Rajvaidya, 1935). Traditional healers used C. dactylon for purifying blood, anuria, biliousness, conjuctivitis, diarrhoea, gonorrhea, itches and stomachache (Muthu et al., 2006). The present study evaluates the effects of dietary administration of C. dactylon ethanolic extract on the potential recovery in C. catla against A. hydrophila, which is associated with haematological, biochemical and histological changes.
2 Material methods
2.1 Fish
The Indian major carp, C. catla, of similar age groups were obtained from the Golden fish farm Karandhai, Thanjavur, Tamil Nadu, India. Average weight of the fishes used for the experiment was 88.05 ± 4.75 g. All fishes were maintained in fiber reinforced plastic (FRP) tanks. The water was replaced once in two days to maintain the water quality. Fishes were kept at the ambient, uncontrolled temperature of 28 ± 2 °C under the natural photoperiod. Fishes were acclimated for 15 days and fed ad libitum with balanced fish diet prepared in the laboratory.
2.2 Bacterial strain
A. hydrophila (ATCC 49140) was cultured in tryptone soya broth (Himedia) for 24 h at 37 °C. The culture broth was centrifuged at 3000g for 10 min. The supernatant was discarded and the pellet was resuspended in phosphate buffered saline (PBS, pH 7.4) for immunization. The optical density (OD) of the bacterial suspension was adjusted to 0.5 and kept in the water bath at 60 °C for 2 h. This bacterial suspension was used for the challenge test. Sterility was confirmed by lack of growth on the nutrient agar. The stock culture in tryptone soya broth was stored at −70 °C with 0.85% (w/v) NaCl and 20% (v/v) to provide stable inocula throughout the study (Yadav et al., 1992).
2.3 Collection of plant and preparation of ethanolic extract of C. dactylon L.
The plant C. dactylon (L.) was collected from Madurai, Tamil Nadu, India, and transferred to PRIST University, Thanjavur, Tamil Nadu, India. It was taxonomically identified and authenticated by Rev. Dr. S. John Britto SJ, Director, The Rapinat Herbarium and Centre for Molecular systematics, St. Joseph College (Autonomous), Thiruchirapalli, Tamil Nadu, India. The voucher specimen was deposited at the Rapinat herbarium and the voucher number is RHCD BP18.
Fresh plant of C. dactylon (L.) was cleaned and shade dried. The dried plants were pulverized by an electrical blender and passed through 20 μm mesh sieve. A powdered plant (550 g) was extracted successfully with 1:2 w/v in 70% ethanol by using soxhlet apparatus. The extraction was carried out in 24 h at room temperature with mild shaking (Chopra et al., 1992). The extract was filtered and concentrated at 45 °C using rotary vacuum evaporator under reduced pressure. The yield of extract was 12.2% w/w in terms of dried starting material.
2.4 Diet preparation and design of experiment
Artificial diet was prepared by using the ingredients fish meal, fish grower, wheat flour, cod liver oil (Universal Medicare Pvt. Ltd.) and vitamin premix (Vetsfarma Ltd.), which contains 20% carbohydrates; 41% proteins; 15% lipids and 9% ash (Table 1). For experimental diet, various concentrations such as 0.05%, 0.5% and 5% level of ethanolic extract of C. dactylon were mixed with the normal diet before pelletization. The control and experimental diet groups were fed with normal diet at the rate of 2% of the body weight per day for 15 days. Experimental feeding was started four weeks prior to immunization.
Ingredients/100 g of Cynodon dactylon extract
Experimental diet (g)
Control diet (g)
Wheat flour
45
45
Dry fish meal
32
32
Fish grower
11
11
Cod liver oil
10
10
Vitamin and mineral premix
2
2
Plant extract (0.05%, 0.5%, 5%)
Present
Absent
Fishes were randomly divided into four groups in triplicate each with 12 fishes (n = 12 fishes/group). They were fed with 0%, 0.05%, 0.5% and 5% (later it will be called as control, group A, group B and group C, respectively) extract of supplemented diet at the rate of 2% of body weight per day for 60 days. Blood was collected for every ten days from each group for assaying particular biochemical parameters.
2.5 Collection of blood samples
For bleeding, each fish was individually caught using a dip net and was bled from the common cardinal vein using 1 ml tuberculin syringe fitted with 24 gauge needle (Michael et al., 1994). In order to collect the blood for serum separation, 200 μl of blood was drawn and whole bleeding procedure was completed within 1 min. The blood was collected in serological tubes and clot was stored at 20 °C overnight. The clot was then spun down at 400g for 10 min. The serum collected by aspiration was stored in sterile eppendorf tubes at −20 °C for further use. After blood collection, the spleen was removed from the fish for analysis of RNA/DNA ratio.
2.6 Haemoglobin content
The haemoglobin content of blood was analysed following the cyanmethaemoglobin method using Drabkins fluid (Qualigens Diagnostics, division of Glaxo Smithkline Pharmaceutical Ltd.). The absorbance was measured using a spectrophotometer (Systronics, UV–Visible spectrophotometer – 118, India) at 540 nm and the final concentration was calculated by comparing with the standard cyanmethaemoglobin (Qualigens Diagnostics). The haemoglobin concentration was calculated by using the following formula: where OD (T) = absorbance of test; OD (S) = absorbance of standard.
2.7 Total erythrocyte and leucocyte count
RBC and WBC diluting fluids were used for taking total erythrocyte count and total leucocyte counts. It was done by mixing 20 μl of blood with 3980 μl of appropriate diluting fluid in a clean test tube and was shaken well to suspend the cells uniformly in the solution. A small drop of this mixture was charged to Neubauer’s counting chamber of haematocytometer and counting was carried out (Larsen and Snieszko, 1964).
2.8 Serum glucose, cholesterol, protein, albumin (A), globulin (G) and A/G ratio
Serum glucose level was determined by Nelson as described by Ashwell (1957); serum cholesterol level was estimated by Lieberman-Bourchard method (Naito, 1985).
Serum protein was estimated by biuret and BCG dye binding method (Reinhold, 1953) using the kit (total protein and albumin kit, Qualigens Diagnostics, Division of Glaxo Smithkline Pharmaceutical Ltd.). Albumin was estimated by bromocresol green binding method (Doumas et al., 1971) with slight modification in micro plate adaptation. The absorbance of standard and test was measured against blank in a spectrophotometer (Systronics, UV–Visible spectrophotometer – 118, India) at 630 nm. Globulin was calculated by subtracting albumin values from total serum protein. A/G ratio was calculated by dividing albumin values and globulin values.
2.9 RNA/DNA ratio
The nucleic acid content of the spleen was examined by the method of Schneider (1957). DNA and RNA were determined using diphenylamine and orcinol, respectively.
2.10 Assay of serum anti-trypsin activity
Serum anti-protease activity was performed and calculated by Vasudeva and Chakrabarti (2004): where A1 = control trypsin activity (without serum); A2 = activity of trypsin remained after addition of serum.
2.11 Preparation of heat killed A. hydrophila bacterin
A. hydrophila was inoculated in tryptone soya broth and incubated overnight in a shaker for 12 h at 28 °C. The culture was centrifuged at 800g for 15 min at 4 °C. The packed cells were washed three times and then the required dose was prepared in phosphate buffered saline (pH 7.2). The bacterial suspension was prepared to 1 × 106 CFU as determined using a Neubauer haemocytometer. The stock cultures in tryptone soya broth (Himedia) were stored at −70 °C with 0.85% (w/v) NaCl and 20% (v/v) to provide stable inocula throughout the study (Yadav et al., 1992).
Heat killed A. hydrophila bacterin was prepared according to the method of Karunasagar et al. (1997).
2.12 Experimental design for specific immunity
Fishes were fed with 0%, 0.05%, 0.5% and 5% (later it will be called as control, group A, group B and group C, respectively) extract of supplemented diet at the rate of 2% of body weight per day for 60 days.
Fishes were randomly divided into four groups (control, group A, group B and group C) in triplicate each with 12 fishes (n = 12 fishes/group) and were fed with experimental diet for 15 days. They were challenged with heat killed A. hydrophila (1 × 106 cells/fish) through intraperitoneal (i.p.) injection. Then, they were fed with the same doses of diet for 7, 14, 21 and 28 days. Total number of fish used in this assay is 144. After the end of every seven days, blood was collected from the fish for assaying specific immunological parameter. Simultaneously, the post-immunized fish spleen of all experimental groups was isolated for histological studies.
2.13 Enzyme linked immunosorbant assay (ELISA)
The fishes were bled at regular intervals of 7 days post immunization to 28 days. Antibody was produced against heat killed A. hydrophila, which were detected using an indirect ELISA according to Binuramesh et al. (2005).
2.14 Plaque forming cell assay (PFC)
Plaque forming cell assay was carried out by the method of Binuramesh et al. (2005, 2006). Six fishes per group were injected i.p. with 0.1 ml of 5% SRBC after 28 days of confinement for evaluating the effect of confinement in all experimental diet groups and control.
2.15 Histological studies
After immunization with A. hydrophila, the experimental fish spleen samples were taken at 7th day and 28th day of feeding trial. Spleen samples were removed and fixed in 10% neutral buffered formalin, embedded in paraffin and stained with hematoxylin and eosin (H&E) for optical examination (Deivasigamani, 2008). Micrograph of spleen was taken from the paraffin sections at a final magnification of 400 using a Nikon Microphot-FXA microscope and an Olympus DP50 camera. For each group, micrographs originating from six fishes (n = 12) were analysed.
2.16 Statistics
The statistical analysis system (SPSS version 17.0) software was used to analyse all the data. The data were expressed as mean ± standard error of mean (SEM) and the data were analysed using the Student’s t-test and one-way analysis of variance (ANOVA) followed by Tukeys post hoc multiple comparison test. Differences were considered statistically significant at P < 0.05 for all the experiments used in this study. All treatments (six fishes in each treatment) were assayed in duplicate for each fish.
3 Result
3.1 Haemoglobin contents, total erythrocyte count and total leucocyte count
Haemoglobin contents and total erythrocyte counts significantly increased in group A, group B and group C at 30–60 days of feeding (Table 2a and b, respectively). Total leucocyte count of C. catla significantly produced higher number of cells from 10 days of feeding. Particularly, leucocyte count was higher in group B and group C than in group A from 40 to 60 days of feeding with the experimental diet (Table 2c).
Days
Control
0.05%
0.5%
5%
(a) Total haemoglobin content
10
6.54 ± 1.02
6.58 ± 1.1
6.61 ± 1.2
6.72 ± 1.43
20
6.55 ± 1.24
6.98 ± 0.1
6.62 ± 1.43
6. 63 ± 0.63
30
6.68 ± 0.3
7.1 ± 1.22*
7.54 ± 0.52*
8.32 ± 0.52*
40
6.75 ± 1.23
9.24 ± 0.9*
9.56 ± 1.05*
10.85 ± 0.7*
50
7.12 ± 1.1
9.11 ± 0.2*
10.58 ± 0.12*
10.86 ± 0.9*
60
7.02 ± 0.12
9.38 ± 0.02*
11.12 ± 0.4*
11.98 ± 1.2*
(b) Total erythrocyte count
10
1.08 ± 0.13
1.18 ± 0.11
1.19 ± 0.07
1.19 ± 0.19
20
1.09 ± 0.11
1.18 ± 0.22
1.21 ± 0.17
1.22 ± 0.01
30
1.19 ± 0.24
1.19 ± 0.32
1.20 ± 0.31*
1.22 ± 0.21*
40
1.2 ± 0.35
1.21 ± 0.07*
1.23 ± 0.21*
1.22 ± 0.38*
50
1.2 ± 0.18
1.22 ± 0.24*
1.22 ± 0.07*
1.23 ± 0.3*
60
1.2 ± 0.07
1.24 ± 0.21*
1.24 ± 0.33*
1.24 ± 0.17*
(c) Total leucocyte count
10
545.11 ± 45.7
608.13 ± 45*
723.18 ± 18.2*
763.13 ± 33.2*
20
548.18 ± 48.7
621.22 ± 32.8*
735.25 ± 19.3*
795.18 ± 41.3*
30
543.21 ± 51.8
645.23 ± 58.1
748.18 ± 17.9*
863.71 ± 12.2*
40
536.15 ± 41.3
670.26 ± 52.6*
767.12 ± 67.4*
888.18 ± 27.2*
50
551.32 ± 50.1
689.15 ± 51.4*
770.77 ± 63.1*
913.12 ± 16.3*
60
552.41 ± 33.1
698.16 ± 13.5*
782.18 ± 12.1*
923.33 ± 7.2*
3.2 Serum glucose and cholesterol
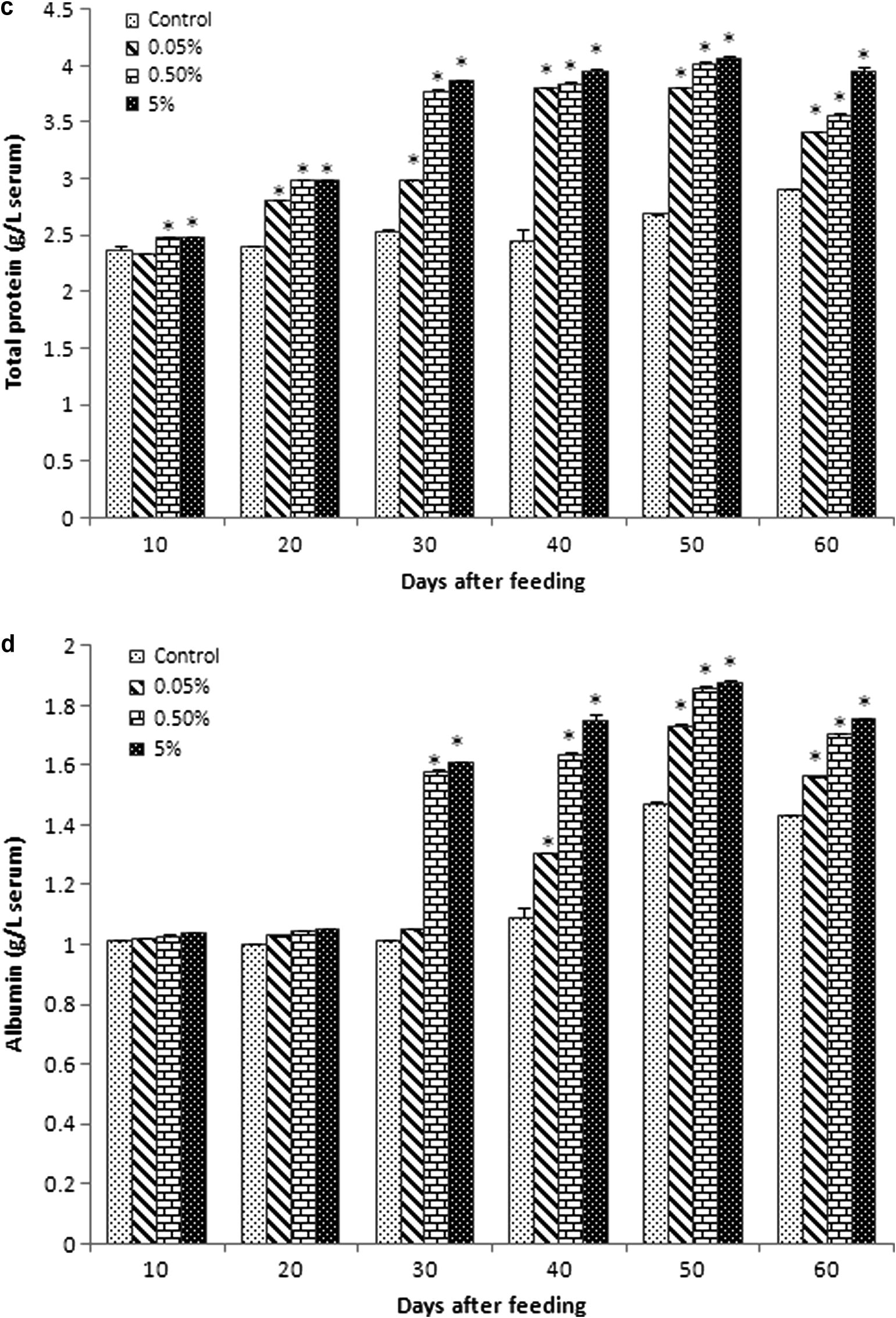
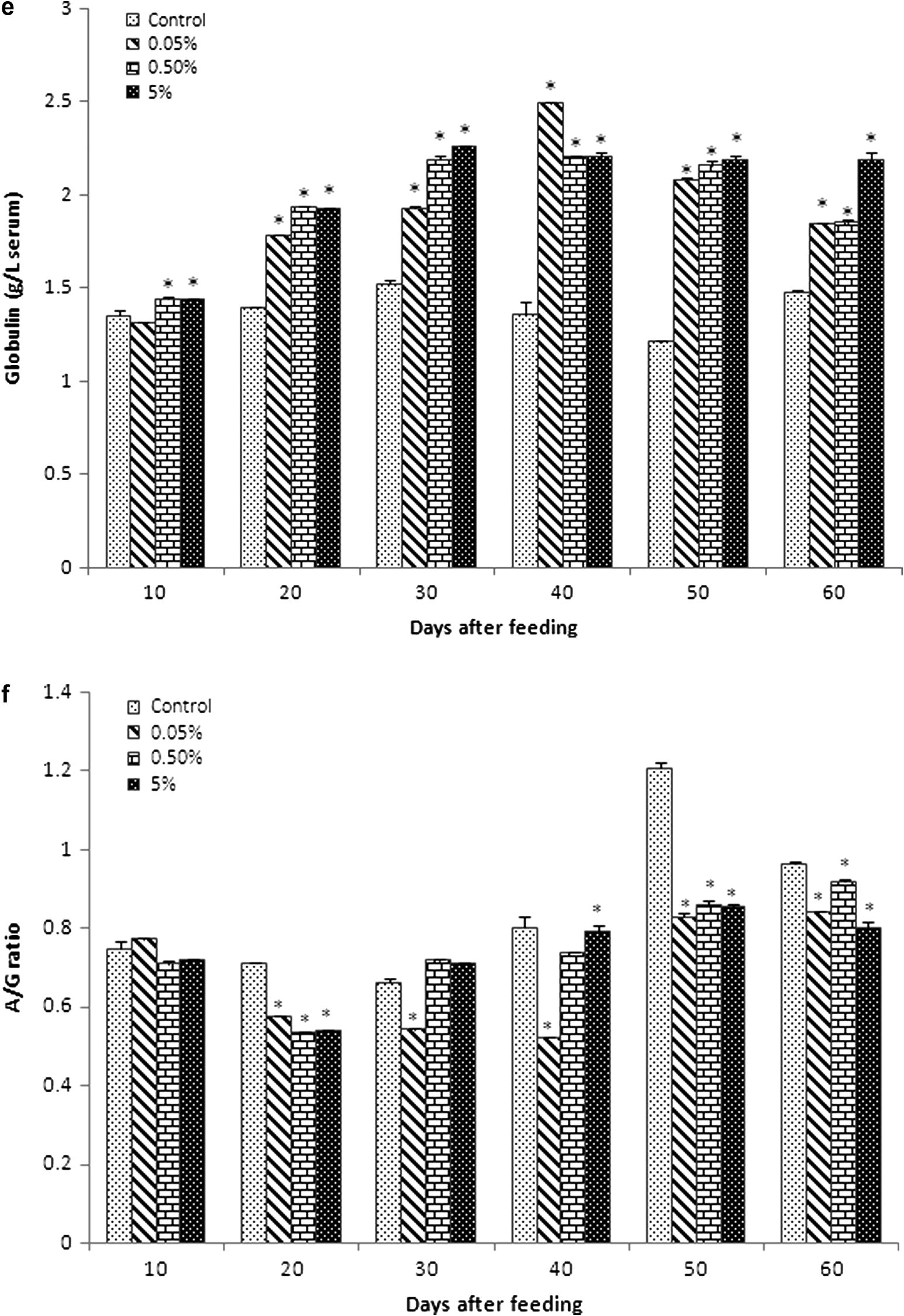
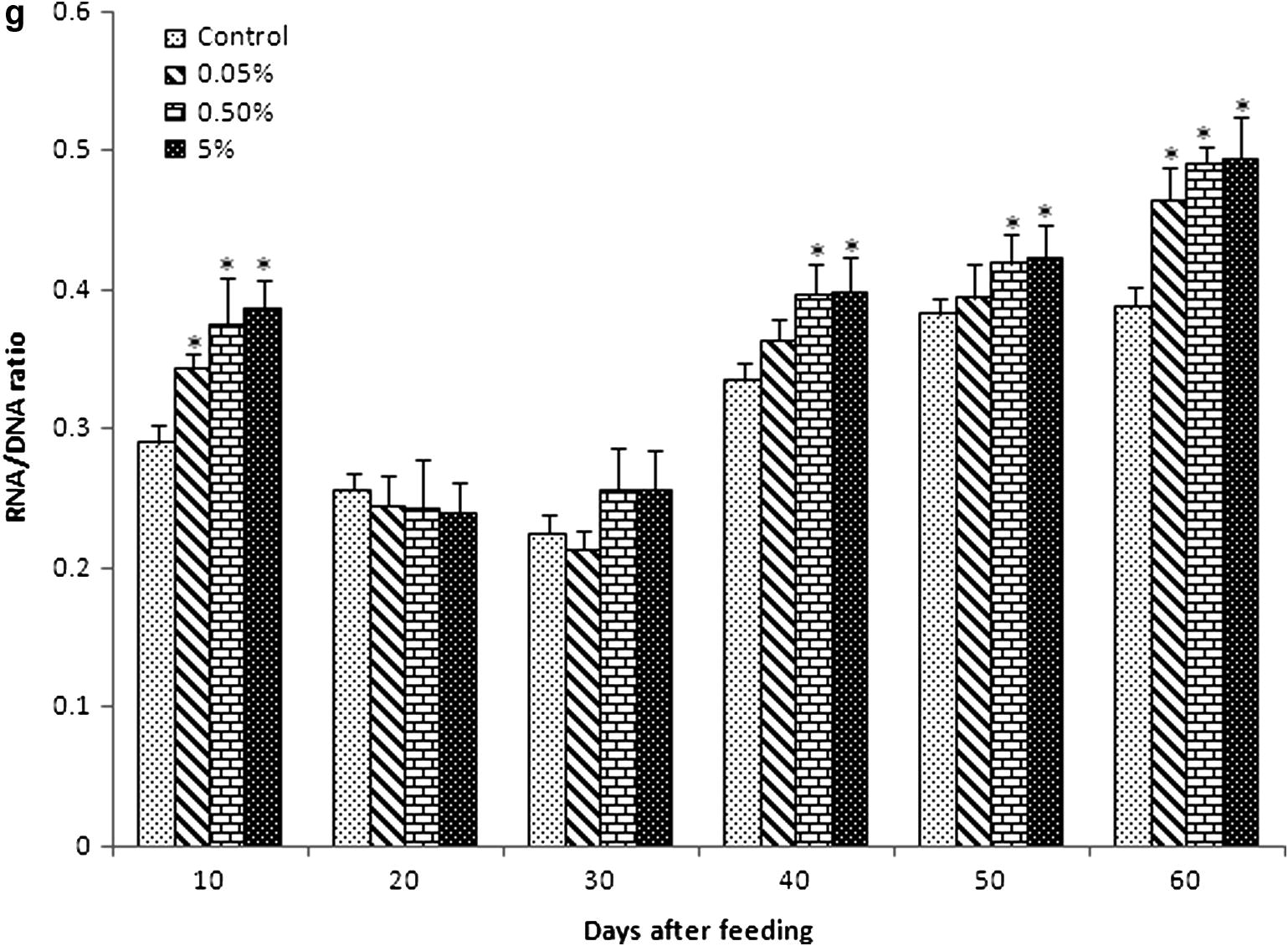
Serum glucose level gradually increased in all experimental diet groups from 10 to 40 days of feeding (Fig. 1a). Glucose levels of group B (423 ± 12.52 and 441.33 ± 10.9) and group C (427.7 ± 21.5 and 445.7 ± 12.33) reached optimum at 30 and 40 days of feeding, respectively. Then, the glucose level slightly reduced at 50 and 60 days, although it was higher than that in the control and group A. In general, control and group A probably showed the same level of glucose.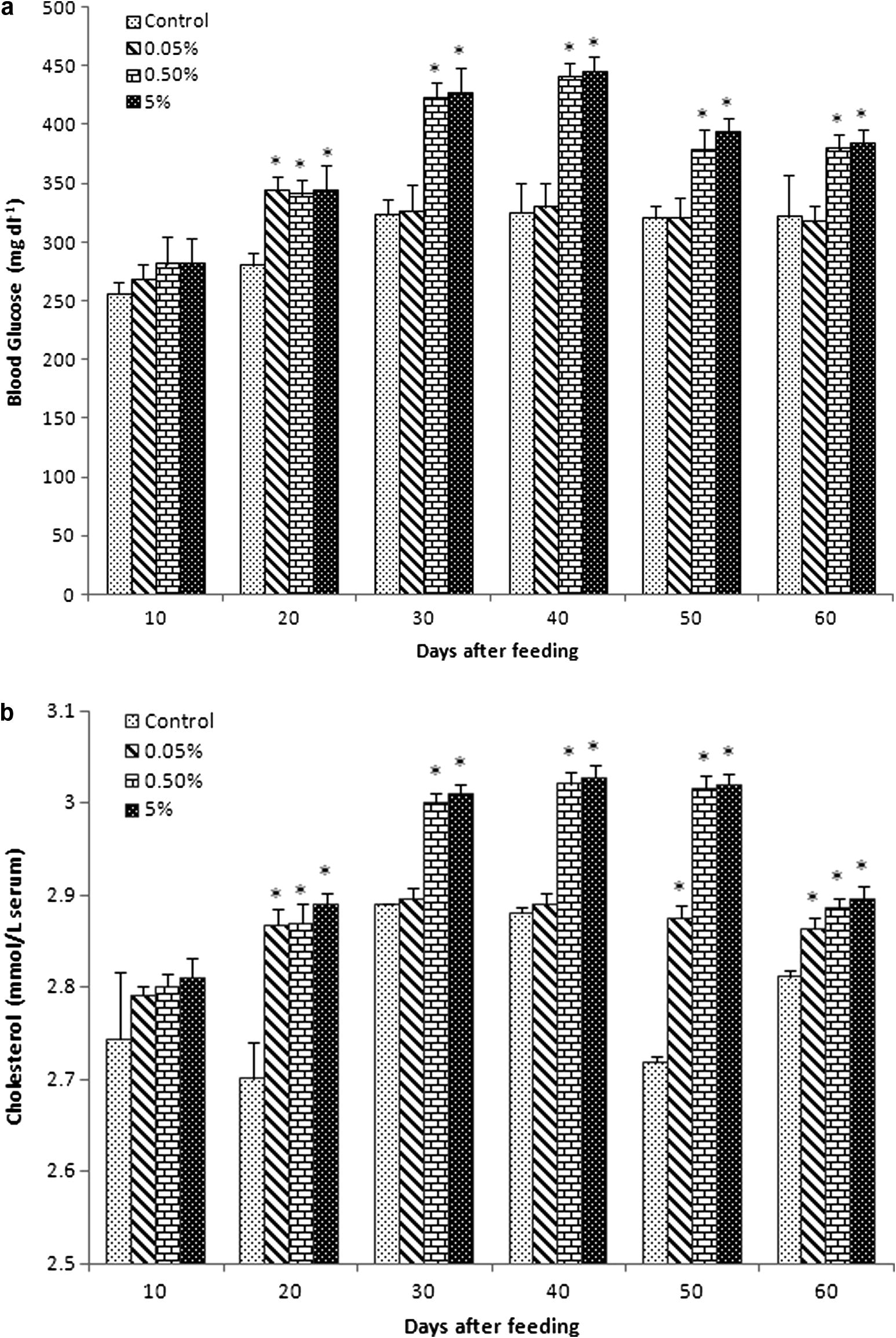
Biochemical values (mean ± SEM) of blood serum in Catla catla fed with different concentrations of C. dactylon mixed diet between normal diet (control). (a) Glucose (Hb: mg dL−1), (b) cholesterol (mmol L−1), (c) total protein (g L−1), (d) albumin (g L−1), (e) globulin (g L−1), (f) albumin/globulin ratio (g L−1), (g) RNA/DNA ratio were obtained at 10, 20, 30, 40, 50 and 60 days of feeding.
Biochemical values (mean ± SEM) of blood serum in Catla catla fed with different concentrations of C. dactylon mixed diet between normal diet (control). (a) Glucose (Hb: mg dL−1), (b) cholesterol (mmol L−1), (c) total protein (g L−1), (d) albumin (g L−1), (e) globulin (g L−1), (f) albumin/globulin ratio (g L−1), (g) RNA/DNA ratio were obtained at 10, 20, 30, 40, 50 and 60 days of feeding.
Biochemical values (mean ± SEM) of blood serum in Catla catla fed with different concentrations of C. dactylon mixed diet between normal diet (control). (a) Glucose (Hb: mg dL−1), (b) cholesterol (mmol L−1), (c) total protein (g L−1), (d) albumin (g L−1), (e) globulin (g L−1), (f) albumin/globulin ratio (g L−1), (g) RNA/DNA ratio were obtained at 10, 20, 30, 40, 50 and 60 days of feeding.
Biochemical values (mean ± SEM) of blood serum in Catla catla fed with different concentrations of C. dactylon mixed diet between normal diet (control). (a) Glucose (Hb: mg dL−1), (b) cholesterol (mmol L−1), (c) total protein (g L−1), (d) albumin (g L−1), (e) globulin (g L−1), (f) albumin/globulin ratio (g L−1), (g) RNA/DNA ratio were obtained at 10, 20, 30, 40, 50 and 60 days of feeding.
Serum cholesterol level of fish was significantly higher than that in the control in all experimental diet groups (Fig. 1b). Cholesterol level of group A was about similar to that of control at 30 and 40 days of feeding (2.89 ± 0.01 and 2.892 ± 0.01), respectively. Cholesterol level was very lesser in control at 20 and 50 days of feeding (2.702 ± 0.04 and 2.72 ± 0.01).
3.3 Serum protein, albumin globulin and A/G ratio
Total protein level increased in all experimental diet groups compared with the control, especially the level of total protein significantly increased from 30 to 60 days of feeding (Fig. 1c).
The albumin level also increased in all experimental groups compared with the control (Fig. 1d). However, the level of albumin in all experimental diet groups was relatively equal to that in the control at 10 and 20 days of feeding. Albumin level immediately raised in group B (1.58 ± 0.004) and group C (1.613 ± 0.001) compared to group A (1.0503 ± 0.00233) and control (1.012 ± 0.002) at 30 days of feeding. Then, gradually increased levels of albumin in group A, group B and group C were seen from 40 to 60 days of feeding compared to the control.
The globulin level progressively increased in all experimental diet groups (Fig. 1e). Level of globulin slightly reduced in group A (1.8543 ± 0.002) and group B (1.8553 ± 0.01) at 60 days compared with 50 days (2.083 ± 0.0311, 2.1623 ± 0.02154, respectively) of feeding. However, group C sustained their level of globulin at 50 (2.1953 ± 0.02) and 60 (2.191 ± 0.04) days of feeding.
The A/G ratio showed a relationship with the increasing percentage of C. dactylon extract (Fig. 1f). Among the control, high difference (1.21 ± 0.013) was found at 50 days of feeding. Significantly, low level of difference was found in group B (0.54 ± 0.002) and group C (0.5431 ± 0.0003) at 20 days feeding; group A showed the values 0.7132 ± 0.002 and 0.7925 ± 0.02 at 30 and 40 days of feeding, respectively.
3.4 RNA/DNA ratio
The RNA/DNA ratio in the spleen of experimental diet group fishes was highest at 60 days of feeding. There was a trend of increase in RNA/DNA ratio correspondingly with the number of days and experimental diet concentration (Fig. 1g). RNA/DNA ratio gradually decreased at 20 and 30 days and then significantly increased at 50–60 days of feeding. On the other hand, RNA/DNA ratio significantly increased at 40–60 days in group C (0.399 ± 0.031, 0.424 ± 0.022, 495 ± 0.031, respectively), when compared with the control (0.335 ± 0.0134, 0.384 ± 0.013, 0.389 ± 0.013, respectively).
3.5 Serum anti-trypsin activity
Serum anti-protease production significantly increased in all experimental diet groups from 10 to 60 days of feeding (Fig. 2). There was no significant enhancement of anti-protease activity observed in group A (69.9 ± 1. 37) compared with control (65.6 ± 1.64) at 10 days of feeding. Among the experimental diet groups, anti-protease activity was higher in group C and low in group A during all experimental periods. Among all the experimental groups, group B was more or less similar to group C except 50 and 60 days of feeding. These results proved that the anti-protease activity directly depends on the concentration of experimental diet.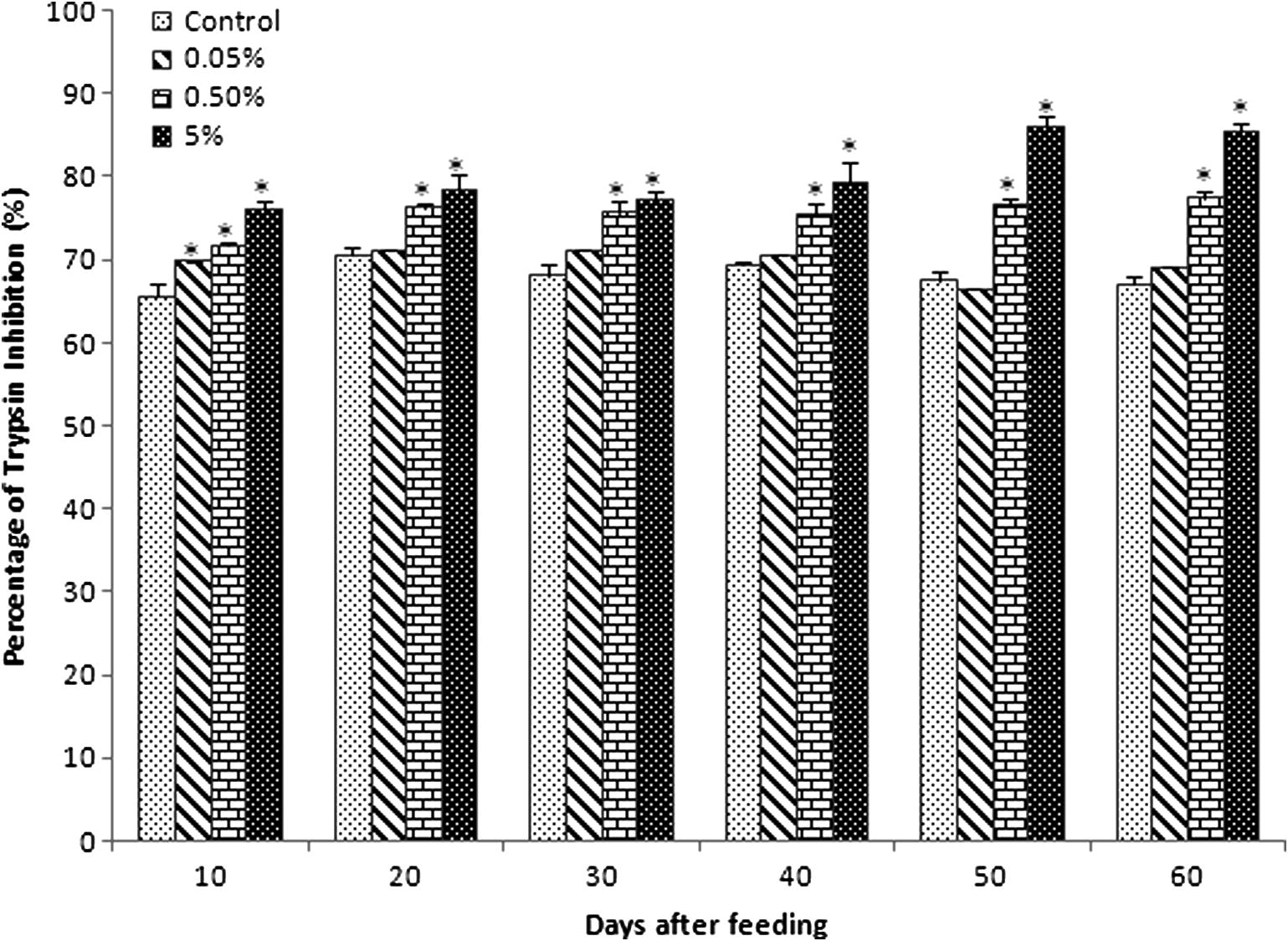
Effect of C. dactylon mixed diet on serum anti-protease activity in C. catla. Each value represents the arithmetic mean ± SEM (*P < 0.05) of six fishes.
3.6 ELISA
ELISA result showed that a significant response in the production of antibody against heat killed A. hydrophila bacterin was observed in group B and group C than in group A and control (Fig. 3). Especially, group B (1.002 ± 0.03) and group C (1.22 ± 0.1) showed optimum response against A. hydrophila at 14 days of feeding and the level of antibody was also higher at 14, 21 and 28 days than group A (0.89 ± 0.01, 0.82 ± 0.001 and 0.65 ± 0.002, respectively) and control (0.89 ± 0.03, 0.81 ± 0.01 and 0.59 ± 0.01, respectively). A minor response of antibody was observed in group A (0.65 ± 0.002) when compared with control (0.59 ± 0.01) against A. hydrophila at 28 days of feeding. Furthermore, the results of group B and group C were more or less similar, which confirmed that the higher concentration of C. dactylon mixed diet provides a better response.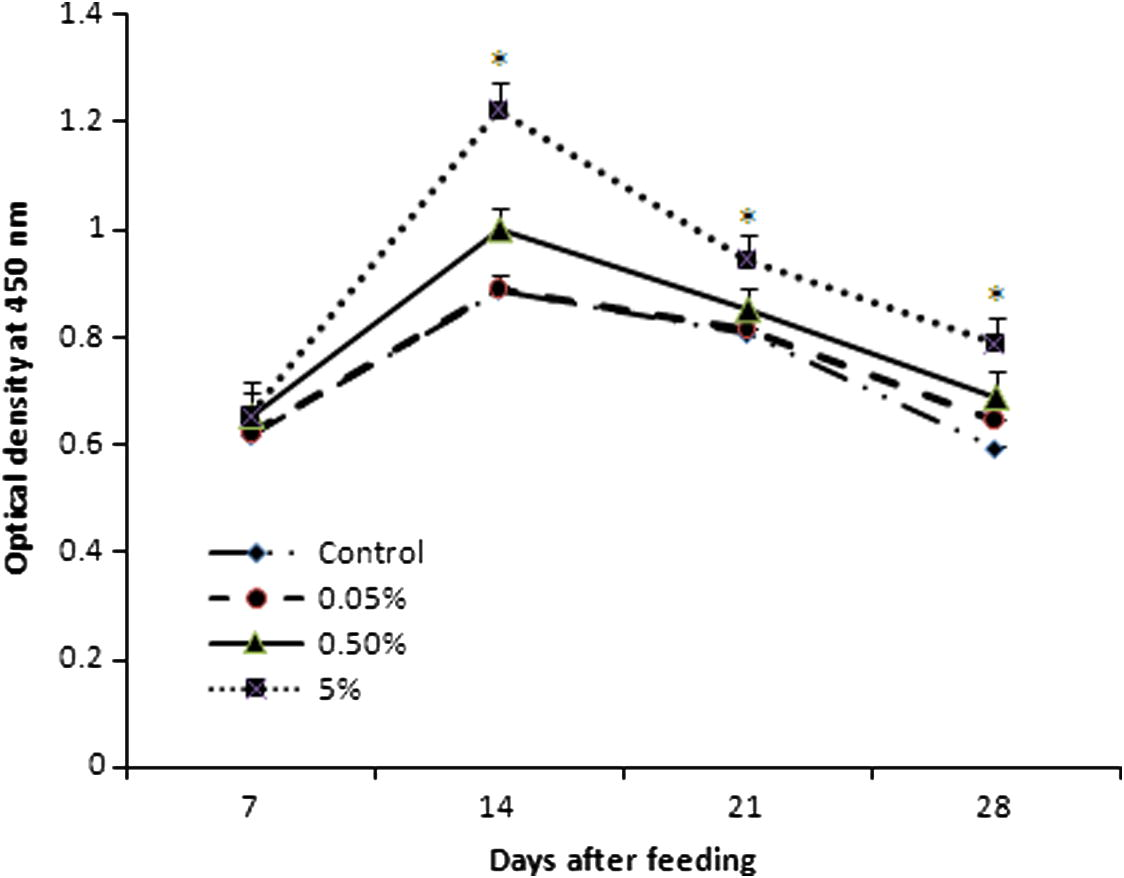
Effect of antibody response to heat killed A. hydrophila in C. catla (n = 6 fishes/group) by ELISA. Each value represents the arithmetic mean ± SEM (*P < 0.05) of six fishes.
3.7 Plaque forming cell assay
The number of antibody producing cells in response to SRBC was observed from 7 to 28 days of feeding in all experimental diet groups (Fig. 4). Among the experimental diet, a higher number of plaque forming cells showed in group C and it reached the maximum (61 ± 4.4) at 21 days of post immunization. The least number of antibody producing cells was observed in control (11 ± 1.5) than group A (38.7 ± 2.33), group B (45 ± 3.4) and group C (47.3 ± 5.9) at 28 days of feeding.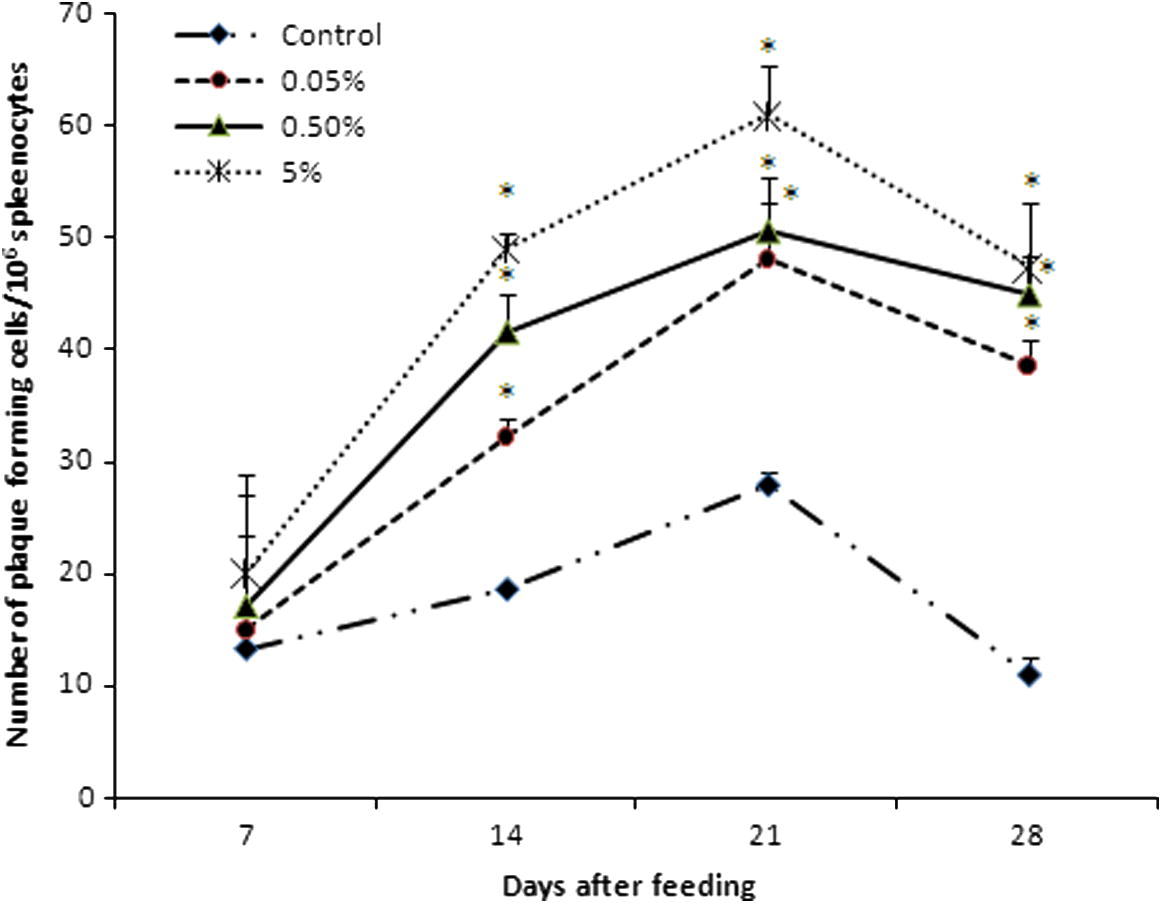
Effect of the number of antibody producing cells against SRBC in C. catla (n = 6 fishes/group) by plaque forming cell assay. Each value represents the arithmetic mean ± SEM (*P < 0.05) of six fishes.
3.8 Histological studies
The spleen of C. catla is a reddish-brown, elongated, thick and flattened structure, lying along the intestine and near the pancreas, trunk-kidney. The spaces within the connective tissue framework are filled with splenic pulp, the distinction of which into red pulp and white pulp is difficult in histological preparations. The pulp in general consists of large venous sinuses and their branches, representing the red pulp (RP), white pulp (WP), nodule-like ellipsoids of lymphoid tissue and dark-staining melanomacrophage centres (MMC) (Fig. 5). Heat killed A. hydrophila challenged fish spleen showed loosely packed red and white pulp (arrow) (Fig. 5a and c); macrophage aggregation can be viewed in Fig. 5b and collapsed MMC and progression of MMC are observed in Fig. 5d–h, which showed that the challenged fishes get immunized and develop their immunological parameters. Through this histological evidence it is proved that C. catla increased their immune response depending on the C. dactylon extract concentration.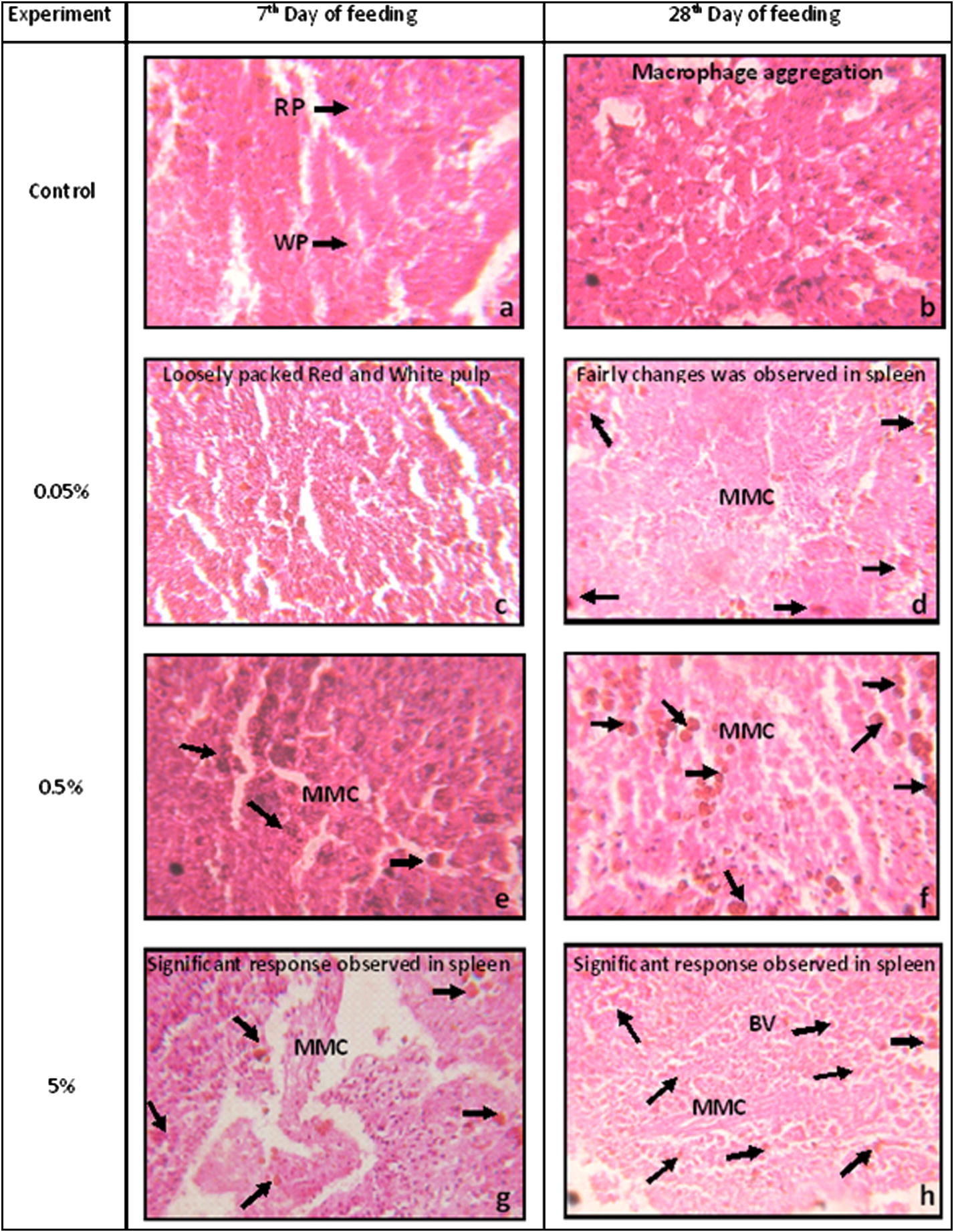
Light micrographs of spleen of Catla catla. Aeromonas hydrophila infected spleen shows loosely packed red and white pulp and collapsed MMC, which shows heavy infection. Arrows: melanomacrophage centres i.e., cluster of macrophage cells. RP: red pulp; WP: white pulp; MMC: melanomacrophage centre; BV: blood vessels.
4 Discussion
The present study was carried out to investigate whether the oral administration of ethanolic extract of plant C. dactylon mixed diet is capable of protecting C. catla against A. hydrophila. Therefore, the plant extract was used for oral administration with prepared diet in the laboratory and to challenge with A. hydrophila for studying the specific immunity on fish. In Indian major carp, outbreak of motile aeromonad septicemia due to the potential opportunistic pathogen A. hydrophila is an easily infectious bacterium, mostly when fishes are exposed to both physical and environmental stressors.
Medicinal plants are progressively being estimated as appropriate alternative sources of antibacterial and antiviral agents. The extract of plant C. dactylon was found to be highly effective in preventing the growth of A. hydrophila infection in C. catla. Similarly, C. dactylon acts as an antibacterial agent against several pathogenic bacteria (Kaleeswaran et al., 2010) and also has antiviral activity against WSSV (White Spot Syndrome Virus) challenged shrimp with no mortality and no signs of WSD (White Spot Disease) at 2% and 40% mortality and 1% in Penaeus monodon (Balasubramanian et al., 2008a). C. dactylon was also found to be highly effective in preventing WSSV infection in black shrimp (Balasubramanian et al., 2008b). The methanolic extract of five different herbal medicinal plants C. dactylon, Aegle marmolos, Tinospora cardifolia, Picrorhiza kurooa and E. alba has shown 74% survival at 800 mg/kg in WSSV-infected shrimp, P. monodon (Citarasu et al., 2006). Similarly, A. hydrophila infected fishes such as Oreochromis mossambicus, Cyprinus carpio, and Labeo rohita were recovered and their survival rate was increased by stimulating the non-specific and specific immune system using O. sanctum (Logambal et al., 2000), E. alba (Christybapita et al., 2007), Mangifera indica (Sahu et al., 2007) and A. aspera (Vasudeva et al., 2006) as feed supplement.
In our study, the total erythrocyte count and haemoglobin level increased from 10 to 60 days of feeding in all the experimental diet groups than in the control. On the other hand, no modulation was observed in the erythrocyte and haemoglobin content of both normal and yeast RNA, ω − 3 fatty acid, β-carotene treated C. catla (Jha et al., 2007). Increasing levels of erythrocyte and haemoglobin content were observed in A. hydrophila infected C. carpio treated with Azadiracta indica (Harikrishnan et al., 2003). The decreased haemoglobin content may be the result of the swelling of RBC as well as poor mobilization of haemoglobin from the spleen and other haemopoietic organs in Ictalurus punctatus (Scott and Rogers, 1981). During the experimental period, the WBC count also increased in the experimental diet groups compared to the control. This result was supported by the study of Sahoo and Mukherjee (1999) who found that WBC count was increased in rohu (L. rohita) fingerlings treated with immunostimulants such as levamisole and ascorbic acid. Jha et al. (2007) also reported that WBC count was increased in C. catla administered with yeast RNA, ω − 3 fatty acid and β-carotene.
In fact, nutritional and immunological conditions are closely related. In the present study, the data showed that the cholesterol level was significantly increased in group B and group C compared to the control, but group A was almost similar to the control at 30 and 40 days of feeding. Glucose is a key product of cellular respiration. Since glucose is an essential fuel for the body tissues, the glucose level was maintained throughout the study period. In the present study, the concentration of serum glucose was increased in C. catla with single exception in group A. Continual increment and then decrement of serum glucose were observed among the control and experimental diet groups. Generally, glucose concentrations were selected as a stress indicator over cortisol or adrenaline level rise in concentration almost immediately following stress. During stressful situation, higher blood glucose is maintained in fish normally through breakdown glycogen from liver, mainly through glycogenolysis (Vijayan et al., 1997). The glucose level increased in the infected or stressed animals (Yoganandhan et al., 2003). It was also reported that the glucose level elevated in stress condition (David et al., 2005). Injection of A. hydrophila was a stress indicator, which was indirectly confirmed by the glucose level during that period and the entire metabolic pathway produces a burst of energy to prepare the fish for an emergency situation (Rottmann et al., 1992). Besides, our result also supported the reduced level of glucose in serum among the control and the experimental diet groups in the normal condition.
Enzyme regulates the addition of glucose to proteins by glycosylation, which is often essential for the body function. Globulins like gamma globulin are essential for maintaining a healthy immune system. Serum albumin and globulin values were always higher in the fish treated with different immunostimulants than those in the control (Choudhury et al., 2005). Increase in the serum protein, albumin and globulin levels is thought to be associated with a stronger innate immune response in fish (Wiegertjes et al., 1996). Our results also increased the serum proteins, albumin, globulin and A/G ratio in all experimental diet groups throughout the experimental days. When compared with the control, serum total protein, albumin and globulin levels were significantly increased in group A, group B and group C, and also a significant difference was found in the anti-protease activity. Thus, the biochemical and immunological parameters also alter the level of anti-protease activity, lysozyme activity, complement factors and serum bactericidal components, which could be correlated with the enhanced leucocytic response and the production of reactive oxygen species (Harikrishnan et al., 2003). As a first line of defense, various peptides such as anti-protease, lysozymes, antibodies, complement factors and other lytic factors are present in serum, where they prevent adherence and colonization of microorganisms (Alexandar and Ingram, 1992) leading to prevention of infection and disease.
Anti-protease contributes to the innate immunity of animals by its bactericidal and anti-inflammatory properties (Jollés and Jollés, 1984). The present study result also revealed that the experimental diet of C. dactylon mixed diet plays a vital role in the enhancement of serum anti-protease activity in all experimental days. This result confirmed that the experimental diet develops the non-specific immunity in C. catla. Similarly, semi purified C. dactylon extract administered to shrimp produced higher level of , which shows the evidence that the plant extract may act as a potential immunostimulant (Balasubramanian et al., 2008b).
This is in an agreement with the observation of Vasudeva and Chakrabarti (2004) that the feeding of C. catla with A. aspera (0.5%) mixed diet for 4 weeks enhanced the level of serum anti-protease level, which might provide resistance against the bacterial pathogens. Similar results also were found in aqueous extract of E. alba which significantly increased the serum anti-protease level after 2 or 3 weeks of feeding in O. mossambicus (Christybapita et al., 2007; Binuramesh et al., 2006). Magnadottir (2006) stated that the anti-protease activity was high in serum by immunization or infection. Correspondingly, C. dactylon mixed diet when fed to C. catla enhanced the protease inhibitor levels. Therefore, the fish can defend more strongly against invading pathogens.
Antibody is the humoral part of the specific immune system to identify and neutralize the foreign objects, followed by invading the pathogens with the help of other humoral and cellular components of the immune system. Enhancement of specific antibody production will facilitate the antigen neutralization and the quick removal from the host. From our results, specific immunity measured by ELISA and plaque forming assay showed higher number of antibodies produced by the experimental diet feeding groups than the control. It proved that the plant extract could develop or induce the specific antibody in fish against the antigen, especially at the higher (5%) concentration. Chakrabarti and Vasudeva (2006) also stated that prophylactic treatment of A. aspera significantly enhanced the specific antibody response and antigen clearance against BSA in C. catla. Vasudeva et al. (2004) and Vasudeva and Chakrabarti (2005a,b) also confirmed that A. aspera enhanced the cRBC-specific hemagglutinating antibody titers and other non-specific immunity in C. catla, L. rohita and C. carpio. Similarly, Spirulina significantly enhanced the antibody titers to keyhole limpet haemocyanin (KLH) in channel catfish I. punctatus. In addition Hemapriya et al. (1997) reported that the intraperitoneal administration of leaf extract of Acalypha indica, Phyllanthus niruri and seed kernel of A. indica enhanced the antibody response in tilapia (O. mossambicus) against sheep red blood cells.
Bacterial entry may be facilitated by injuries. C. catla do not have protective scales and their fins can easily scratch the skin of another fish within the immediate vicinity. A. hydrophila infected C. catla showed the symptoms such as haemorrhages, erosion of the pectoral fins, ulcers on the skin surface and septicemia. After 48 h of infection in the present investigation, the following symptoms were observed in C. catla: fluid in the abdomen, swollen liver, spleen and kidney with clear histopathological alterations in morbid fish. Moreover, histopathological changes have been widely used as biomarkers for evaluating the health of fish. It was also stated that histopathological alterations are good biomarkers for field assessment (Alagappan et al., 2009). Yambot and Inglis (1994) described that the infection of A. hydrophila caused acute mortality among Nile tilapia in which the most apparent clinical signs included opaqueness in one or both eyes, accompanied by exophthalmia and eventual bursting of the orbit. Deivasigamani (2008) stated that humoral and cell mediated immunity have the ability to endeavor immune response against sheep red blood corpuscles (SRBC) in catfish. Mortality due to typical histopathological lesions, necrosis of sheathed arteries in the spleen has occurred. In addition to changes in MMC, it is relatively easy to measure and they could have valued as non-specific and specific biomarkers (Wolke, 1992; Bols et al., 2001). Our study also revealed that the spleen was loosely packed with white and red pulp in control group compared to the experimental diet groups of C. catla. Aggregation of MMC was higher in the experimental diet group than the control and considerable changes were seen in the histology of spleen tissue. It is an evidence for the ability of C. dactylon extract used for controlling the Aeromonas outbreak.
In conclusion, our study has shown that experimental diet treatment at 5% level might have potential efficacy on immunity in C. catla. C. dactylon mixed diet is able to stimulate the biochemical and immune parameters in C. catla. These results suggested that C. dactylon extract may provide a new therapeutic value in specific and non-specific immunity in the fresh water fish C. catla. However, the effect of C. dactylon mixed diet used as an immunostimulant by oral delivery has to be further investigated in relation to molecular mechanism to evaluate its efficacy of disease resistance. Necessary field trials should be adapted in fish culture practices. In addition, sanitation and good management practice will reduce Aeromonas outbreak in a fish production unit.
Acknowledgements
Financial and excellent supports from PRIST University, Thanjavur, Tamil Nadu, India, and the technical guidance given by Dr. R. Dinakaran Michael are gratefully acknowledged.
References
- Histopathological alterations in estuarine catfish (Arius maculatus; Thunberg, 1792) due to Aeronomas hydrophila. Infect. World J. Fish Mar. Sci.. 2009;1:185-189.
- [Google Scholar]
- Noncellular non-specific defense mechanisms of fish. Annu. Rev. Fish Dis.. 1992;2:249-279.
- [Google Scholar]
- Bacterial Fish Pathogens: Disease in Farmed and Wild Fish. West Sussex, England: Ellis Horwood Ltd.; 1987. pp. 13–350
- Studies on the immunomodulatory effect of extract of Cyanodon dactylon in shrimp, Penaeus monodon, and its efficacy to protect the shrimp from white spot syndrome virus (WSSV) Fish Shellfish Immunol.. 2008;25:820-828.
- [Google Scholar]
- Oral administration of antiviral plant extract of Cynodon dactylon on a large scale production against white spot syndrome virus (WSSV) in Penaeus monodon. Aquaculture. 2008;279:2-5.
- [Google Scholar]
- Effect of chronic confinement stress on the immune responses in different sex ratio groups of Oreochromis mossambicus (Peters) Aquaculture. 2005;250:47-59.
- [Google Scholar]
- Effect of sex ratio on the immune system of Oreochromis mossambicus (Peters) Brain Behav. Immun.. 2006;20:300-308.
- [Google Scholar]
- Achyranthes aspera stimulates the immunity and enhances the antigen clearance in Catla catla. Int. Immunopharmacol.. 2006;6:782-790.
- [Google Scholar]
- Glossary of Indian Medicinal Plants. New Delhi: CSIR Publication; 1992. p. 50
- Dietary yeast RNA supplementation reduces mortality by Aeromonas hydrophila in rohu (Labeo rohita L.) juveniles. Fish Shellfish Immunol.. 2005;19:281-291.
- [Google Scholar]
- Oral administration of Eclipta alba leaf aqueous extract enhances the non specific immune responses and disease resistance of Oreochromis mossambicus. Fish Shellfish Immunol.. 2007;23:840-852.
- [Google Scholar]
- Influence of selected Indian immunostimulant herbs against white spot syndrome virus (WSSV) infection in black shrimp, Penaes monodon with reference to haematological, biochemical and immunological changes. Fish Shellfish Immunol.. 2006;21:372-384.
- [Google Scholar]
- Blood glucose and glycogen levels as indicators of stress in the freshwater fish, Labeo rohita under fenvalerate intoxication. J. Ecotoxicol. Environ. Monit.. 2005;15:1-5.
- [Google Scholar]
- Enhancement of nonspecific immunity and disease resistance in Oreochromis mossambicus by Solanum trilobatum leaf fractions. Fish Shellfish Immunol.. 2007;23:249-259.
- [Google Scholar]
- J. Fish Dis.. 1998;21:317-320.
- Albumin standards and measurement of serum albumin with bromocresol green. Clin. Chim. Acta. 1971;31:87-96.
- [Google Scholar]
- Some medicinal plants as immunostimulant for fish. J. Ethnopharmacol.. 2003;88:99-106.
- [Google Scholar]
- Hematological and biochemical parameters in common carp, Cyprinus carpio, following herbal treatment for Aeromonas hydrophila infection. Aquaculture. 2003;221:41-50.
- [Google Scholar]
- Immunostimulation by leaf extracts of a few south Indian medicinal plants in Oreochromis mossambicus (Peters) Dev. Comp. Immunol.. 1997;21:175.
- [Google Scholar]
- Haemato immunological responses to dietary yeast RNA, ω − 3 fatty acid and b-carotene in Catla catla juveniles. Fish Shellfish Immunol.. 2007;23:917-927.
- [Google Scholar]
- Screening of phytochemical properties and antibacterial activity of Cynodon dactylon L. Int. J. Curr. Res.. 2010;3:83-88.
- [Google Scholar]
- Immunization with bacterial antigens: infection with motile Aeromonas. Dev. Biol. Stand.. 1997;90:135-141.
- [Google Scholar]
- Resistance to Vibrio alginolyticus in juvenile rockfish (Sebastes schlegeli) fed diets containing different doses of aloe. Aquaculture. 1999;180:13-21.
- [Google Scholar]
- Effect of dietary carbohydrate on haematology, respiratory burst activity and histological changes in L. rohita juveniles. Fish Shellfish Immunol.. 2005;19:331-344.
- [Google Scholar]
- Modification of the microhematocrit technique with trout blood. Trans. Am. Fish Soc.. 1964;90:139-142.
- [Google Scholar]
- Immunostimulatory effect of leaf extract of Ocimum sanctum Linn. in Oreochromis mossambicus (Peters) Hydrobiologia. 2000;430:113-120.
- [Google Scholar]
- A rapid method for repetitive bleeding in fish. Indian J. Exp. Biol.. 1994;32:838-839.
- [Google Scholar]
- Medicinal plants used by traditional healers in Kancheepuram District of Tamil Nadu, India. J. Ethanobiol. Ethnomed.. 2006;2:1-10.
- [Google Scholar]
- Cholesterol: review of methods, check sample PTS 85-1. Chikago: American Society of Clinical Pathology; 1985. pp. 1–17
- Effects of dietary lipids histological changes in hepatic tissues of Catla catla fingerlings. Indian J. Vet. Pathol.. 2004;28:121-124.
- [Google Scholar]
- The use of immunostimulants to increase resistance of aquatic organisms to microbial infections. In: Shariff M., Subasinghe R.P., Arthur J.R., eds. Disease in Asian Aquaculture. Proceedings of the First Symposium on Diseases in Asian Aquaculture. Vol vol. 1. Philippines: Asian Fisheries Society; 1992. p. :39-50.
- [Google Scholar]
- Abdominal dropsy disease in major carps of Meghalaya: isolation and characterization of Aeromonas hydrophila. Curr. Sci.. 2005;88:1897-1900.
- [Google Scholar]
- Rajvaidya, S.G., 1935. Shankar Nighantu, Banousadhi Bhander, Jabalpur, MP, India.
- Manual determination of serum total protein, albumin and globulin fractions by biuret method. In: Reiner M., ed. Standard Method of Clinical Chemistry. New York: Academic Press; 1953. p. :88.
- [Google Scholar]
- Modulation of the non-specific defence of fish by structurally conserved microbial polymers. Fish Shellfish Immunol.. 1999;9:269-290.
- [Google Scholar]
- Rottmann, R. W., Francis-Floyd, R., Durborow, R., 1992. The Role of Stress in Fish Disease. SRAC Publication No. 474, p. 4.
- Influence of the immunostimulant chitosan on immune response of healthy and cortisol-treated rohu (Labeo rohita) J. Aquac. Trop.. 1999;14:209-215.
- [Google Scholar]
- Effect of Magnifera indica kernel as a feed additive on immunity and resistance to Aeromonas hydrophila in Labeo rohita fingerlings. Fish Shellfish Immunol.. 2007;23:109-118.
- [Google Scholar]
- Determination of nucleic acids in tissues by pentose analysis. In: Colowick S.P., Kaplan N.O., eds. Methods in Enzymology. Vol vol. 3. Academic Press; 1957. p. :680-682.
- [Google Scholar]
- Hematological effects of prolonged sublethal hypoxia on channel catfish Ictalurus punctatus (Rafinesque) J. Fish Biol.. 1981;18:591-601.
- [Google Scholar]
- Enhanced antiproteases in Labeo rohita fed with diet containing herbal ingredients. Ind. J. Clin. Biochem.. 2004;19:132-134.
- [Google Scholar]
- Stimulation of immunity in Indian major carp Catla catla with herbal feed ingredients. Fish Shellfish Immunol.. 2005;18:327-334.
- [Google Scholar]
- Dietary incorporation of Achyranthes aspera seed influences the immunity of common carp Cyprinus carpio. Indian J. Anim. Sci.. 2005;75:1097-1102.
- [Google Scholar]
- Potentiation of antibody production in Indian major carp Labeo rohita, rohu, by Achyranthes aspera as a herbal feed ingredient. Aquaculture. 2004;238:67-73.
- [Google Scholar]
- Effect of Achyranthes aspera on the immunity and survival of infected with Aeromonas hydrophila. Fish Shellfish Immunol.. 2006;20:263-273.
- [Google Scholar]
- Metabolic responses associated with confinement stress in tilapia: the role of cortisol. Comp. Biochem. Physiol.. 1997;116:89-95.
- [Google Scholar]
- Immunogenetics of disease resistance in fish; a comparable approach. Dev. Comp. Immunol.. 1996;20:365-381.
- [Google Scholar]
- Fish diseases and disorders: viral, bacterial and fungal infections. CABI. 1998;3:479-511.
- [Google Scholar]
- Cytotoxin elaboration by Aeromonas hydrophila isolated from fish with epizootic ulcerative syndrome. J. Fish Dis.. 1992;159:183-189.
- [Google Scholar]
- Yambot, A.V., Inglis, V., 1994. Aeromonas hydrophila isolated from Nile tilapia (Oreochromis niloticus) with “eye disease”. In: International Symposium on Aquatic Animal Health, University of California, School of Veterinary Medicine, Davis, CA. p. 103.
- Effect of two Chinese herbs (Astragalus radix and Scutellaria radix) on non specific immune response of tilapia, Oreochromis niloticus. Aquaculture. 2006;253:39-47.
- [Google Scholar]
- Biochemical physiological and hematological changes in white spot syndrome virus-infected shrimp, Penaeus indicus. Aquaculture. 2003;221:1-11.
- [Google Scholar]




