

Translate this page into:
Are owl pellets good estimators of prey abundance?
⁎Corresponding author at: Centro Nacional Patagónico (CENPAT-CONICET), Boulevard Almirante Brown 2915 (U9120ACD), Puerto Madryn, Chubut, Argentina. Tel.: +54 0280 488 3184. andrade@cenpat-conicet.gob.ar (Analia Andrade),
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Some ecologists have been skeptics about the use of owl pellets to estimate small mammal’s fauna. This is due to the assumptions required by this method: (a) that owls hunt at random, and (b) that pellets represent a random sample from the environment. We performed statistical analysis to test these assumptions and to assess the effectiveness of Barn owl pellets as a useful estimator of field abundances of its preys. We used samples collected in the arid Extra-Andean Patagonia along an altitudinal environmental gradient from lower Monte ecoregion to upper Patagonian steppe ecoregion, with a mid-elevation ecotone. To test if owls hunt at random, we estimated expected pellet frequency by creating a distribution of random pellets, which we compared with data using a simulated chi-square. To test if pellets represent a random sample from the environment, differences between ecoregions were evaluated by PERMANOVAs with Bray–Curtis dissimilarities. We did not find evidence that owls foraged non-randomly. Therefore, we can assume that the proportions of the small mammal’s species in the diet are representative of the proportions of the species in their communities. Only Monte is different from other ecoregions. The ecotone samples are grouped with those of Patagonian steppes. There are no real differences between localities in the small mammal’s abundances in each of these ecoregions and/or Barn owl pellets cannot detect patterns at a smaller spatial scale. Therefore, we have no evidence to invalidate the use of owl pellets at an ecoregional scale.
Keywords
Tyto alba
Pellet contents
Prey abundance
Random hunt
Northern Patagonia

1 Introduction
Oliver Pearson, a pioneer in Patagonian mammalogy, always said that owls were his best field assistants during Patagonian surveys. They hunted more species and more individuals than his trap lines, so they were useful estimators of field abundance. However, estimation of small mammal’s abundance is difficult and controversial. Different methods can be used but each of these has some limitations and produces special biases on the abundance assessments. Trap success is a technique frequently used, where the trap-attracted species are over-estimated and trap-shy are under-estimated. Besides, the efficiency of trapping is related with the ecological habits of the small mammal species surveyed (ground-dwelling, scansorial, fossorial, etc.), requiring a different type of trap sampling. The environmental heterogeneity and specificity of small mammal habitat selection demands large sampling effort to properly assess an area. This can be possible in studies on species population dynamics or community composition analysis; but difficult or even impossible at landscape scale surveys (for advantages and biases of different sample techniques, see Millán de la Peña et al., 2003; Torre et al., 2004).
A complementary method is the analysis of raptor pellets. Owls swallow their prey as a whole and expel undigested remains, such as bones, compacted in hair and feathers. Ecologists and mammalogists have been skeptical about the validity of owl pellets as samples representing relative abundance of prey species in an area. The use of pellet contents as an estimate of relative abundance of small mammal in the field depends on two assumptions: (a) owls hunt at random, and (b) pellets represent a random sample of species ingested (Yom-Tov and Wool, 1997). Despite its frequent use for different ecological purposes, controversy still persists about the fact that whether abundances of each prey in the owl’s diet represent the proportion of species in the field. Some studies reported a positive association between trapping data and pellet sampling (Bernard et al., 2010; Glue, 1971; Hanney, 1962; Mikkola, 1983; Terry, 2010) while others did not find such association (Perrin, 1982; Torre et al., 2004). Avenant (2005) demonstrated that Barn owls are efficient samplers of the small mammals because they can detect more species and can more accurately sample the species abundance compared to trapping exercised over the same period. Terry (2010) found a high fidelity of the death assemblages to the living community in terms of richness, evenness, taxonomic composition, and rank and proportional abundances of the prey species.
Although some studies compared the small mammal abundances data collected by two complementary methods (owl pellet and trapping), yet even if abundance estimations are different, it does not exclude pellets as appropriate methodology in species relative abundance studies. Both methods have their own biases; therefore we believe that it is wrong to use one of these (traps) to validate the other (pellets sample). This is especially relevant with Barn owl (Tyto alba) pellets, as these were employed to measure biodiversity in several ecological studies (Avery et al., 2002, 2005; Bernard et al., 2010; González-Fischer et al., 2012; Heywood and Pavey, 2002; Lyman, 2012; McDowell and Medlin, 2009a; Millán de la Peña et al., 2003; Torre et al., 2004). Fidelity of Barn owls for the same roosting place produces large amounts of small mammal bones that accumulate over time and become a part of sediments. This particular habit enables paleontologists to employ these assemblages in taphonomical, paleoecological and paleoenvironmental reconstructions, based on the variations in the proportions of small mammal’s species over time (Andrews, 1990; Avery, 2001; Fernández-Jalvo, 1995; Pearson, 1987; Pearson and Pearson, 1993; Teta et al., 2005; etc.).
In Argentinean Patagonia, food habits of Barn owls were assessed along latitudinal (Trejo and Lambertucci, 2007) and also elevational gradients (Travaini et al., 1997). Both studies concluded that Barn owl pellets are a good complement to trapping in attempts to efficiently sample a small mammal community. Moreover, Travaini et al. (loc. cit.) concluded that Barn owl diets reflected the real composition of cricetid rodents along the altitudinal gradient and that consumption of prey species was dependent on their availability. Recently, a study was undertaken at a landscape scale to document the effects of altitudinal gradients on the community composition, abundance, and species richness of small mammals in Patagonian arid lands using Barn owl pellets (Andrade and Monjeau, 2014). The study reflected spatial variation of community composition of small mammals along the altitude gradient. Pellet samples were also employed to define geographic distribution ranges of some small mammal species (Andrade, 2008; Martin, 2003; Nabte et al., 2009; Udrizar Sauthier et al., 2007, 2008, 2011).
Barn owl (T. alba) is a medium sized, active hunter owl, and territorial in its feeding habitat. It is mainly nocturnal but may also be active during the early evening and morning hours. Its hunting area may vary from 400–500 m to 2–3 km, depending on food availability, and it searches for prey by silently flying over open areas. It feeds on vertebrates, mainly rodents (Andrews, 1990). McDowell and Medlin (2009b) and Heywood and Pavey (2002) showed that Barn owls can switch to alternative prey only when rodents are scarce and may return to preferred preys as soon as they become available.
Yom-Tov and Wool (1997) provides the only study which indirectly tests that pellets constitute a random sample of prey small mammal species. The study worked with the hypothesis that pellets are a random sample of their catch. The study concluded that Barn owls are though not selective hunters yet the contents of the pellets could be biased toward larger preys. Andrews (1990) evaluated the response of Barn owl to prey size and found that the more abundant small mammal species in the field was more abundantly consumed by the owl, though it adapts to different sized prey depending on their availability. Bernard et al. (2010) found that Barn owl is an opportunistic predator, though density/availability of other prey species can affect their relative consumption.
The objectives of this paper were to test the two assumptions in the use of pellet contents as an estimate of relative abundance of small mammal in the field and to address two derived questions: (1) do Barn owls hunt at random? and (2) are pellet contents of this owl a useful tool to characterize the small mammal’s assemblages sampled in different habitats at the arid Patagonia? We also discuss the advantages and biases in the estimation of rodent abundances through the pellet content analysis.
2 Material and methods
2.1 Study area
Continental Patagonia, located in the southernmost end of South America (between 39°S and 55°S) presents a sharp environmental gradient, due to the synergy between precipitation and temperature (Paruelo et al., 1998). Gradually decreasing precipitation sets boundaries of the main vegetation types from west to east: forest, bunchgrass steppe, brush-grass steppe and bush steppe (León et al., 1998). Extra-Andean arid Patagonia is located east of the Andean mountains, lying between sea level and 1800 m above sea line (a.s.l). This sharp elevational gradient is product of diverse basalt flows. The climate is arid, warmer in the eastern and northern faces and lower elevations and colder in the southern and western faces and higher elevations. Western and southern faces exhibit a wide temperature range; from −25 °C (winter, June–September) to above 35 °C (summer, December–March) and average annual precipitation is 187 mm (Maquinchao Meteorological Station, data given by DPA – Departamento Provincial de Aguas – Río Negro). For the east, average annual temperature and precipitation are 14 °C and 227 mm, respectively (Dique 1 Meteorological Station, Valcheta, Río Negro, data given by DPA). Precipitation is concentrated during winter. Wind direction in the area is generally from the west or south–west to east or north–east with average velocity of 3.15 m/s (Maquinchao station). Some localities remain under snow or with frosts even in late spring (September).
Northern Extra-Andean arid Patagonia comprises three major ecoregions or phytogeographical provinces, from west to east: the Patagonian steppe (cold desert), the Monte (Southern Monte, warm desert) and a broad ecotonal area in between (Ecotono Rionegrino) (León et al., 1998). A rich small mammal fauna inhabits these southern latitudes. At least 20 small mammal species (including rodent families, Cricetidae, Caviidae and Ctenomyidae, and Didelphidae oposums) have been reported along the elevation gradient, characterized by volcanic plateaus of northern Extra-Andean Patagonia (Andrade, 2009; Andrade and Monjeau, 2014). A species turnover exists over the elevation gradients. Species typical of the warmer Monte desert disappear and give place to a typical colder Patagonian desert fauna around 1000 m a.s.l. As Fig. 1 shows, the altitude ranges of most of the species overlap in a vast ecotone area at the intermediate levels (between 750 and 1000 m a.s.l.) in which the vegetation is typical of the Ecotono Rionegrino (Andrade, 2009; Andrade and Monjeau, 2014).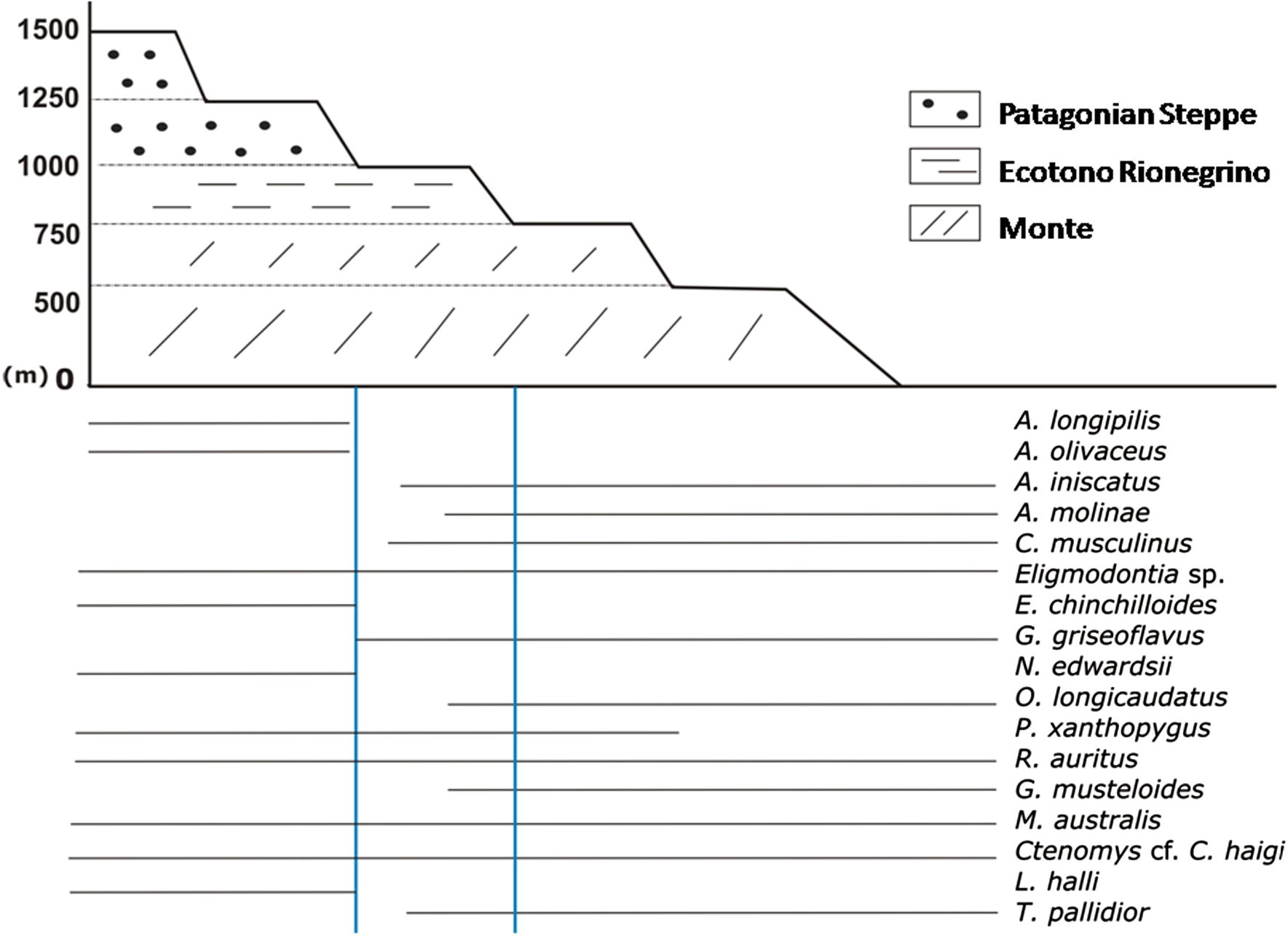
Scheme that represents the distribution of small mammal species and the approximate boundaries of ecoregions along the elevational gradient from northern Extra-Andean Patagonia.
2.2 Study samples and data analysis
Owl pellet samples were collected at 3 ecoregions from northern Extra-Andean Patagonia: Southern Monte, Patagonian steppes and Ecotono Rionegrino, following the elevation gradient. Abundances of small mammal’s species in those samples have been presented in Table 1. Prey items were identified to the lowest taxonomic level using reference collections and identification keys (Andrade, 2009). The number of prey items was estimated by counts of the jaw and crania remains.
Species
PL
EL I
EL II
CM I
CM II
P I
P II
P III
M I
M II
SC
ER
C I
SP
M I
M II
T I
T II
SC
SM
SM
SM
SM
SM
ER
ER
ER
PS
PS
PS
PS
PS
PS
PS
PS
PS
PS
PS
A. longipilis
2
1
1
19
2
6
1
7
7
5
9
5
6
A. olivaceous
42
24
15
62
42
41
8
31
18
59
19
61
10
54
A. iniscatus
13
7
5
2
55
10
4
3
A. molinae
10
15
21
4
59
C. musculinus
136
1
12
1
13
3
C. macronyx
2
1
6
2
9
5
Eligmodontia sp.
86
83
21
2
49
80
27
28
153
68
52
11
19
12
79
40
48
11
55
E. chinchilloides
93
26
13
65
49
204
13
63
31
75
47
129
94
13
G. griseoflavus
33
15
28
16
95
5
2
1
3
2
L. micropus
N. edwardsii
5
3
2
30
26
10
4
3
1
1
1
O. longicaudatus
6
2
13
P. xanthopygus
5
6
8
75
69
32
38
17
6
15
2
7
5
11
4
15
10
5
R. auritus
8
1
7
6
8
28
15
4
39
33
36
20
3
3
51
9
28
32
5
Ctenomys sp.
62
9
7
3
1
13
9
17
20
19
14
2
7
33
15
32
31
G. leucoblephara
1
2
4
37
6
4
4
M. australis
5
17
5
83
2
7
2
2
2
1
1
4
3
1
L. halli
4
3
8
7
5
5
5
4
2
1
3
5
T. pallidior
1
24
3
4
61
14
4
2
Total
349
166
135
57
549
376
161
113
415
257
389
75
130
90
332
144
335
203
144
To test the first assumption (owls forage at random), we looked for an association between prey compositions in different pellets. If owls forage at random each pellet is a random sample from the environment. Thus, the difference among pellets should also be random. Therefore, an association χ2 comparing expected (under the null hypothesis of random foraging) and real prey frequencies in pellets should not be significant. We used a randomization procedure to approximate χ2 expected values. This procedure was necessary because some expected values were zero. Pellets are combinations of preys; so many possibilities are mathematically plausible. As some combinations were not present in our pellet sample, their frequencies, and consequently, their expected values were zero. Randomization creates the expected values for all combinations of species. For this purpose, 107 random prey compositions were drawn using a two-step procedure. First, we assigned a size (number of preys) to each composition. We sampled this value from prey numbers in pellets. Second, we sampled the species at random to fill the compositions for the assigned size. The probability of selecting a species was proportional to its abundance in pellets. All samples were random and with replacement. The number of times each composition appeared in the total 107 random prey compositions was the expected value. We estimated p-values using the simulation procedure described in R (R core team, 2014). We used 5000 simulations to approximate each p-value.
Three pellet samples were employed for these association χ2 analysis (Table 1): Puerto Lobos (PL, Southern Monte, 122 pellets, 42° 00′ 003″ S, 65° 04′ 19″ W, Udrizar Sauthier and Pardiñas, 2006), Sierra Chacays (SC, Patagonian steppes, 60 pellets, 42° 48′ 31″ S, 68° 00′ 56″ W), and Arroyo Pinturas (P III, Ecotono Rionegrino, 37 pellets, 41° 42′ 31″ S, 66° 43′ 50″ W).
To test de second assumption (pellets represent a random sample from the environment), differences among ecoregions in the prey composition were calculated using a PERMANOVA (Anderson, 2008). Differences were calculated using the Bray–Curtis dissimilarity index. We performed four PERMANOVAs, one with all ecoregions included and one for each pair of them (Monte/Steppe; Monte/Ecotone; Steppe/Ecotone).
3 Results
Statistical analysis allowed us to conclude that in the arid Patagonia, Barn owls hunt at random. Non-significant values of association chi-square (Monte: χ2 = 2911.90; p = 0.4658; Ecotono Rionegrino: χ2 = 553.48; p = 0.8236; Patagonian steppes: χ2 = 303.69; p = 0.6277) indicate that expected frequency values (calculated under the null hypothesis of random foraging) and real prey frequencies in pellets are similar. In other words, no pellet deviated from the catch distribution in any of the environments, favouring our hypothesis that owls hunt at random. Therefore owl sampling can be an acceptable method for sampling small mammal abundances in an area.
We can also accept the second assumption: pellets represent a random sample from the environment. Owl pellets can be used to discriminate different environments at the scale of ecoregions, but cannot be used to discriminate more refined environmental subdivisions. PERMANOVA indicates significant differences between ecoregions (F3,16 = 5.4365, p = 0.001; Fig. 2), though differences are non-significant when Monte ecoregion was excluded (PERMANOVA without Ecotono Rionegrino: F2,13 = 9.0338, p = 0.002; PERMANOVA without Patagonian steppes: F1,5 = 3.1894, p = 0.038; PERMANOVA without Monte: F2,13 = 2.2291, p = 0.098, Fig. 2). In other words, there are no significant differences between samples collected in the Patagonian steppes and the Ecotono Rionegrino. We conclude that owl consumption can be used to separate environments, but only if this difference is substantial, which is likely to occur at the ecoregional scale. This result also implies that owls adapt their variation in its diet under local differences in small mammal abundance.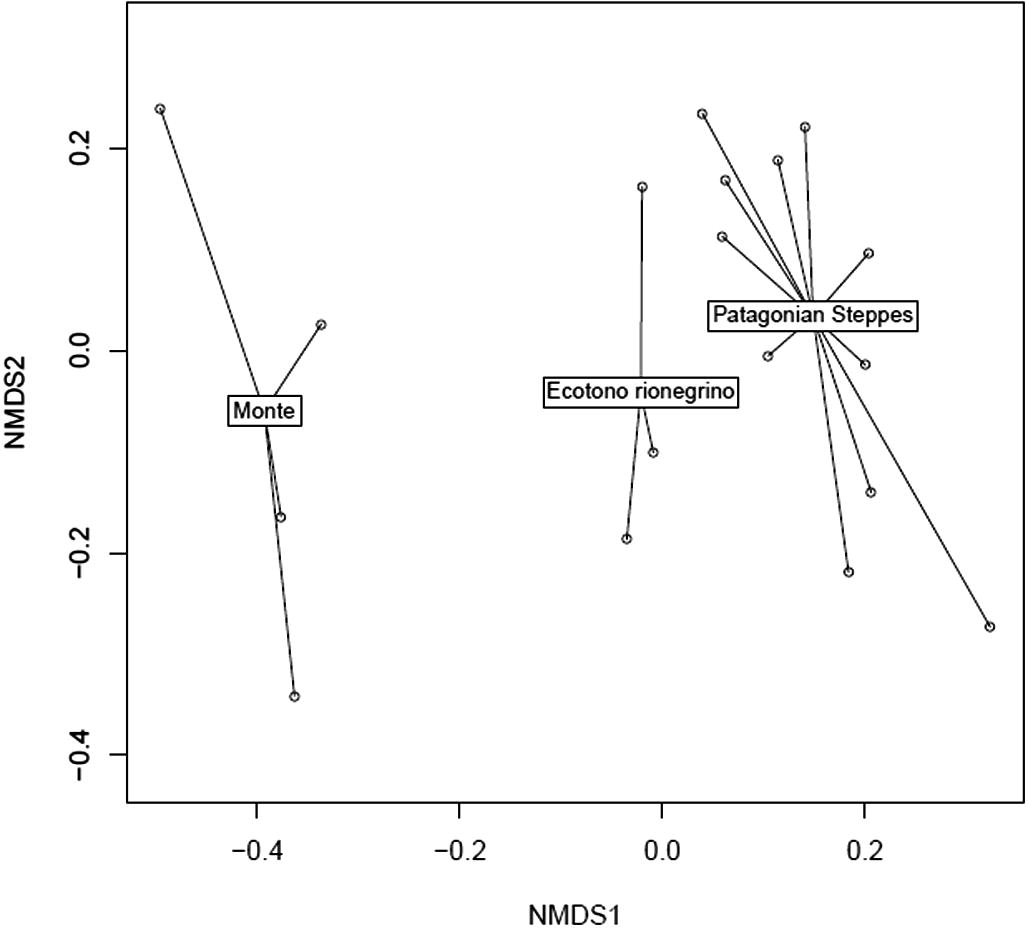
Non-metric multidimensional scaling (NMDS) of prey abundance in owl pellets. Note that there is a strong separation between Monte and the other ecoregions, which is also reflected in our PERMANOVAs (NMDS stress: 0.09584).
4 Discussion
Our results let us to prove that Barn owls hunt at random. This allows us to answer the first question: Are owls useful field assistants? We think so. Barn owl captures preys randomly in a non-selectively way on the landscape. Therefore, we can state that it consumes the species in similar proportion as they are in the field. This supports the study of Andrews (1990) reporting that Barn owls pellets preserve the original community equitability, consuming more of the common species. The relevance of Barn owl pellets in biodiversity assessments has been indicated in previous studies exhibiting a higher species richness compared with that of trap samplings performed in the same area (Andrade and Monjeau, 2014; Andrews, 1990; Avenant, 2005; Torre et al., 2004).
Some limitations of owl pellets as surrogates of abundance can be mentioned. First, there are certain limits to owls’ ability to transport prey, much like other aerial predators that hunt while flying. Andrews (1990) pointed out that Barn owls can adapt to different sized prey depending on availability and size diversity in the small mammal’s community. However, an upper limit was estimated at 200–250 g, with an optimal prey weight between 80 and 100 g (Andrews, 1990; Dodson and Wexlar, 1979; Yom-Tov and Wool, 1997). A bias toward larger prey must be taken into consideration when using pellets to estimate prey abundances.
Second, the activity pattern of the predator may also bias the abundance estimation based on owl pellets. Barn owls are mainly nocturnal, so pellet contents could underestimate diurnal prey species. In the arid Patagonia, those were represented by species of the families Caviidae and Ctenomiidae (Galea leucoblephara, Microcavia australis and Ctenomys sp., Table 1).
Third, hunting strategy is another feature to be considered. Barn owls hunt in open areas, searching for prey by silent flight and taken them mostly on the ground (Marti, 1974). This situation makes pellet contents as a suitable methodology for abundance estimations on open areas, such as the one considered in our study, but inadequate to sample communities in forests, dense brushy habits and ecotones between open and densely covered landscape.
These conclusions led us to answer the second question: Are pellet contents of this owl a useful tool to characterize the small mammal’s assemblages sampled in different habitats at the arid Patagonia? Pellets represent a random sample from the environment, but an “owl pixel” resolution is needed to be defined because some limitations arise concerning the geographic scale. Barn owl pellets are a suitable methodology to characterize small mammals’ communities from different ecoregions. PERMANOVA suggest significant differences between the Monte and the Patagonian steppes, but no significant differences between samples collected inside each ecoregion. In line with previous studies (Andrade and Monjeau, 2014), different small mammals assemblages characterized the Monte, Patagonian steppes and ecotone area (Ecotono Rionegrino). However, the ecotone samples were grouped with Patagonian steppes. Some implications could be derived from this proposition. The most important one is related to owl hunting area and the spatial scale employed in the study. Barn owls explore its territory in a radius of 400 m–3 km around its perch depending on food availability (Andrews, 1990). Considering the highest surface in poor environments and assuming hunting territory as circular (28.27 km2), we can define the resolution of the space unit we can coin as the “owl pixel”. The size of the owl pixel is a limitation to characterize differences in small mammal composition at the habitat and microhabitat scales. At larger spatial scale analysis, this limitation is only relevant if the environment inside the owl pixel is not homogeneous in the availability of habitat types for the small mammals species considered. In the arid Extra-Andean Patagonia, the assumption of homogeneity is quite reasonable, especially for the Monte and Patagonian steppes, two extensive ecoregions. In those open and homogeneous areas, owl pellet contents are good estimators of small mammal abundance. However, this issue may come into play in sharp environmental gradients (such as the elevation gradient) or narrow ecotonal zones, where owls may be sampling between different elevations and/or environments. This resolution issue could answer why ecotone samples were grouped altogether with those of Patagonian steppes. The spatial scale employed in the analysis and the homogeneity of habitat types inside the owl pixel must be considered in biodiversity assessments that employed pellet contents as a sampling technique, to allow those to provide accurate measurements.
Finally, based on our results and the discussion above, we can affirm that Barn owl pellet samples are a suitable methodology in biodiversity assessments taking into account the biases and limitations discussed here.
5 Conclusions
This study tested an important assumption necessary to consider when using Barn owl pellets to estimate small mammal’s fauna: owls hunt their prey in a non-selective way. From this, it follows that Barn owl pellets’ contents are a useful method to estimate relative abundances of its preys. Proportions of the small mammals in the diet are representative of the real proportions of the species in their communities. Taking into consideration some of the biases imprinted by this kind of data source –underestimation of diurnal and larger prey-, and ensuring habitat homogeneity inside the owl resolution (an area of 28.27 km2), pellets are a suitable methodology in biodiversity assessments at an ecoregional scale in open habitats.
References
- A new method for non-parametric multivariate analysis of variance. Aust. Ecol.. 2008;26:32-46.
- [Google Scholar]
- Mammalia, Rodentia, Cricetidae, Notiomys edwadsii (Thomas, 1890): distribution extension and geographic distribution map. Checklist. 2008;4(1):33-36.
- [Google Scholar]
- Andrade, A., 2009. Ecología Geográfica y Biodiversidad de los Pequeños Mamíferos de la Meseta de Somuncurá (Provincias de Río Negro y del Chubut) (Ph.D. thesis). Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires.
- Patterns in community assemblage and species richness of small mammals across an altitudinal gradient in semi-arid Patagonia, Argentina. J. Arid Environ.. 2014;106:18-26.
- [Google Scholar]
- Owls, Caves and Fossils: Predation, Preservation, and Accumulation of Small Mammal Bones in Caves, with an Analysis of the Pleistocene Cave Faunas from Westbury-Sub-Mendip, Somerset, UK. Chicago: University of Chicago Press; 1990.
- Barn owl pellets: a useful tool for monitoring small mammal communities? Belg. J. Zool.. 2005;135(supplement):39-43.
- [Google Scholar]
- The Plio-Pleistocene vegetation and climate of Sterkfontein and Swartkrans, South Africa, based on micromammals. J. Hum. Evol.. 2001;41:113-132.
- [Google Scholar]
- Micromammalian distribution and abundance in the western Cape Province, South Africa, as evidenced by Barn owls Tyto alba (Scopoli) J. Nat. Hist.. 2005;39(22):2047-2071.
- [Google Scholar]
- A contribution from Barn owl pellets to known micromammalian distributions in KwaZulu-Natal South Africa. J. Afr. Zool.. 2002;37(2):131-140.
- [Google Scholar]
- Dietary response of Barn Owls (Tyto alba) to large variations in populations of common voles (Microtus arvalis) and European water voles (Arvicola terrestris) Can. J. Zool.. 2010;88:416-426.
- [Google Scholar]
- Small mammal taphonomy at La Trinchera de Atapuerca (Burgos, Spain). A remarkable example of taphonomic criteria used for stratigraphic correlations and palaeoenvironment interpretations. Palaeogeogr. Palaeoclimatol. Palaeoecol.. 1995;114:167-195.
- [Google Scholar]
- Seasonal variations in small mammal-landscape associations in temperate agroecosystems: a study case in Buenos Aires province, central Argentina. Mammalia. 2012;76:399-406.
- [Google Scholar]
- Observation on the food of the Barn Owl in southern Nyasaland and a method of ascertaining population dynamics of rodent prey. Ann. Mag. Nat. Hist.. 1962;6:705-713.
- [Google Scholar]
- Relative importance of plague rodents and dasyurids as prey of Barn owls in central Australia. Wildl. Res.. 2002;29:203-207.
- [Google Scholar]
- Grandes unidades de vegetación de la Patagonia extra andina. Ecol. Aust.. 1998;8:125-144.
- [Google Scholar]
- Rodent-prey content in long-term samples of barn owl (Tyto alba) pellets from the Northwestern United States reflects local agricultural change. Am. Midl. Nat.. 2012;167:150-163.
- [Google Scholar]
- Nuevas localidades para marsupiales patagónicos (Marsupialia: Didelphimorphia y Microbiotheria) en el Noroeste de la Provincia del Chubut. Mastozool. Neotrop.. 2003;10:148-153.
- [Google Scholar]
- Using the diet of the barn owl (Tyto alba) as an indicator of small vertebrate abundance in the Channel Country, south-western Queensland. Aust. Mamm.. 2009;31:75-80.
- [Google Scholar]
- The effects of drought on prey selection of the barn owl (Tyto alba) in the Strzelecki Regional Reserve, north-eastern South Australia. Aust. Mamm.. 2009;31:47-55.
- [Google Scholar]
- Owls of Europe. Poyser, London: T & A.D; 1983.
- Response of small mammal community to changes in western French agricultural landscapes. Landscape Ecol.. 2003;18:265-278.
- [Google Scholar]
- Mammalia, Rodentia, Sigmodontinae, Akodon molinae (Contreras, 1968): new locality records and filling gaps. Checklist. 2009;5(2):320-324.
- [Google Scholar]
- The climate of Patagonia: general patterns and controls on biotic processes. Ecol. Aust.. 1998;8:85-101.
- [Google Scholar]
- Mice and the postglacial history of the Traful Valley of Argentina. J. Mamm.. 1987;68:469-478.
- [Google Scholar]
- La fauna de mamíferos pequeños de Cueva Traful I, Argentina: pasado y presente. Præhistoria. 1993;1:211-224.
- [Google Scholar]
- Prey specificity of the Barn Owl in the Great Fish River valley of the eastern Cape Province. S. Afr. J. Wildl. Res.. 1982;12:14-25.
- [Google Scholar]
- On raptors and rodents: testing the ecological fidelity and spatiotemporal resolution of cave death assemblages. Paleobiology. 2010;36:137-160.
- [Google Scholar]
- Micromamíferos (Didelphimorphia y Rodentia) y paleoambientes del Holoceno tardío en la Patagonia noroccidental extra-andina (Argentina) Archaeofauna. 2005;14:183-197.
- [Google Scholar]
- Three methods for assessing richness and composition of small mammal communities. J. Mamm.. 2004;85(3):524-530.
- [Google Scholar]
- Food habits of Common Barn Owls along elevational gradient in Andean Argentine Patagonia. J. Raptor Res.. 1997;31:59-64.
- [Google Scholar]
- Feeding habits of Barn Owls along a vegetative gradient in northern Patagonia. J. Raptor Res.. 2007;41:277-287.
- [Google Scholar]
- Mammalia, Marsupialia, Didelphidae, Lestodelphyshalli: new records, distribution extension and filling gaps. Checklist. 2007;3:137-140.
- [Google Scholar]
- Enlarging the knowledge on Graomys griseoflavus (Rodentia: Sigmodontinae) in Patagonia: distribution and environments. Mammalia. 2011;75:185-193.
- [Google Scholar]
- Micromamíferos terrestres de Puerto Lobos, Chubut, Argentina. Mastozool. Neotrop.. 2006;13(2):259-262.
- [Google Scholar]
- Mammalia, rodentia, sigmodontinae, loxodontomys micropus: new locality records. Checklist. 2008;4:171-173.
- [Google Scholar]
- Do the contents of barn owl pellets accurately represent the proportion of prey species in the field? Condor. 1997;99:972-976.
- [Google Scholar]




