

Translate this page into:
Ultra Violet (UV-B) radiation intrudes Cadra cautella reproductive biology by influencing vitellogenin expression
⁎Corresponding author.
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Peer review under responsibility of King Saud University.
Abstract
Objective
Date palm is a very important fruit crop growing in the world's warm climate. Several pests attack date fruit in the field and in storage. The almond moth, Cadra cautella, is a primary key pest which damages date fruits. Present research shows ultraviolet irradiation effect on mortality and reproduction of C. cautella female larval and female pupal stages in the laboratory.
Methods
Female larva aged 18 days and female pupae aged 1–2 days, were directly exposed to ultra violet (UV) type B in a chamber for 0, 2, 4, 6, and 12 h. Mortality of treated larvae and pupae was calculated and compared with the 0 h treatment (control). To quantify vitellogenin gene expression through real time polymerase chain reaction (RT-qPCR), larvae and pupae samples were collected from each treatment after each exposure interval and preserved at −80 °C.
Results
Cadra cautella larvae and pupae, were vulnerable to the UV and the mortality was observed for all exposure intervals showing the highest mortality rate of 90 % larval mortality at 12 exposure interval. Whereas, in case of pupal mortality, 72.26 % mortality was observed after 12 h of exposure intervals. The UV-treated female pupae that survived the pupal developmental stage and emerged as adult moths were not healthy and were mostly deformed, preventing them from emerging from the puparium. Furthermore, when treated female moths emerged, they were unable to copulate with normal male moths, resulting in fewer unfertile eggs. In addition, the lethal effects of UV on vitellogenin gene expression in treated larvae and pupae were confirmed using real-time PCR, the results confirmed that UV had a harmful effect on vitellogenin gene expression.
Conclusions
The current study results are highly encouraging and may be useful in suppressing C. cautella populations in storages and managing this renowned pest of stored date fruits and other agricultural commodities on an industrial/commercial scale.
Keywords
Date fruit
Storage
Cadra cautella
Saudi Arabia
Ultraviolet

- UV
-
Ultra Violet
- PCR
-
polymerase chain reaction
- RNAi
-
RNA interference
- PBS
-
Phosphate Buffered Saline
Abbreviations
1 Introduction
Food security is becoming increasingly important, and the need of agricultural food commodities that can be preserved after harvesting is critical. Pest damage and agricultural commodity loss in storage and retail stores are critical. Growing grains/pulses crops and bringing food from field to table is a risky, expensive, and time-consuming process which is well understood by the farmers who are actively involved in crop production. Severe infestations of this insect on date fruit have a negative impact on date fruit quality, aesthetic value, and trade. The almond moth, Cadra cautella (Lepidoptera, pyralidae) may attack numerous agricultural commodities during storage, including wheat, cereals, legumes, and date fruits. The insect is cosmopolitan and can completely destroy the item that has been attacked if no timely control measures are taken (Aldawood et al., 2013, Husain et al., 2017, Jian, 2019). Its generation grows very fast, female produce several hundred eggs, larvae eats and damage the produce. In dates, the infestation starts in the field when the date fruit is partially dry but still present on a bunches attached to the tree (Aldawood, 2013). Whereas, in the storage, cracks and crevices are the safest places where pest can hide and start infestation immediately after dates arrived in storage.
A variety of attempts are being made to inhibit the pest population, but unfortunately the issue has so far emerged and is rising continuously, increasing the risk of commercial trade. For dry-consumable agricultural commodities, fumigation with methyl bromide was the best option, but its use was either prohibited or reduced due to human health and environmental risks from chemicals (Bulathsinghala and Shaw, 2014). Scientists are trying to find technologies and methods that are environmentally safe, cost-effective and user-friendly, to handle the stored consumer insects, like C. cautella using modified and controlled atmospheres (Navarro, 2012; Li and Zhang, 2015; Yan et al., 2016; Husain et al., 2017), use of ozone gas (Husain et al., 2015), entomapathogenic fungi, essential oils, ultraviolet and gamma radiations (Alwaneen et al., 2019), and the cultural practices to combat the situation. Therefore, in the present study, we required to see that ultraviolet light might affect the vitellogenin gene expression and reproductive potential in C. cautella.
In oviparous species, vitellogenin is critical for the production of yolk protein (vitellin) (Oie et al., 1975; Sappington, 2002; Tufail et al., 2014; Husain et al., 2021) Vitellogenin is produced in animal fat tissues and transferred to the developing embryo, where it acts as food for the developing oocytes (Tufail and Takeda, 2002; Shu et al., 2009). Vitellogenin has been sequenced, characterized, and silenced through RNA interference (RNAi) in several insects including C. cautella (Yano et al., 1994; Tufail et al., 2007; Veerana et al., 2014; Moriyama et al., 2016; Husain et al., 2021; Rasool et al., 2021). The Vg gene was chosen due to its role in insect reproduction, as it has been reported that it contributes 88–93 % of the yolk protein in the yellow fever mosquito, Aedes aegypti, and the Germen cockroach, Blattella germanica, respectively (Hagedorn and Judson, 1972; Oie et al., 1975). Full-grown C. cautella larvae leave the feeding arena and roam around looking for a suitable location to pupate (Husain et al., 2017). This larval activity gives us the opportunity to use some technique that may affect the physiology of the pupae or adult. Use of ultraviolet radiation in this regard could be a better opportunity to interrupt physiological functions, as pupae are fixed on the outer surfaces of food products.
Many insect species associated with stored agricultural products as well as household stuff and field crop pests have been observed with radiation effects (Faruki et al., 2007; Heidari et al., 2016). Gamma radiation were used for apple decontamination and accepted as quarantine approach for safe transportation of dried food products. Ultraviolet radiation has proved very lethal for different insect species associated with stored products, their effect has been documented from interference of insect's genetic material (DNA) to the mortality (Hori et al., 2014). Arthropods usually pass through several stages of metamorphosis during their development time and the UV have potential to terminate their physiology through interrupting the ecdysone hormone which is very important for both metamorphosis and insect reproduction.
We intended to investigate the expression of the vitellogenin gene affected by UV radiation in C. cautella adult female larvae and pupae. The study comprises of the following particular objectives. Investigating the ultraviolet radiation effect on C. cautella adult female larvae and pupae survival, examining the ultraviolet radiation effect on vitellogenin expression in female moth, as well as studying the influence of ultraviolet radiation on reproduction of adult female moth after being exposed to UV radiation.
2 Materials and methods
2.1 Cadra cautella culture rearing
Since 2009, Cadra cautella colony has been raised in our laboratory on an artificial diet using poultry diet elements as primary components (Aldawood et al., 2013). In our laboratory, more than a hundred generations of C. cautella have been reared using the conventional approach for handling and growing this insect. The C. cautella colony is susceptible because we did not transfer any field populations into the main colony or subject it to any harmful treatment/factor that could lead to population evolution or resistance.
At emergence, male and female moths were collected in a plastic container where they mate and lay eggs. The plastic box is specifically constructed for this purpose, with a mesh under the lid through which eggs pass and aggregate in the plastic box's lid cover, where they can be collected. The collected eggs are placed in another properly labeled plastic box with an artificial diet. The egg laying dates are confirmed on the box label. These boxes are placed in controlled environments where eggs hatch and larvae continue to develop.
2.2 Female larvae and pupae collection
At 25 °C, eggs hatch in 3–4 days (Husain et al., 2017). The immature larvae start to eat the diet and molt within few days to develop to the next instar. The 4th molting occurs on the 15th day of age, and when the larvae enter the 5th instar, the reproductive organs begin to develop, and the testes in male larvae become visible on the 18th day of age. So, at 20 days of the age, the female larvae were collected from these boxes and placed separately in a plastic cup of 50 g capacity. The female larvae were given 0.25 g of fresh made artificial diet. After a few days, the female larvae were grown into pupae and employed in the UV bioassay. The pupae were 1–2 days old at the time of the UV bioassay.
2.3 Ultraviolet irradiation application to C. cautella female larvae and pupa
In this experiment, two 15-watt UV-B lamps (Wisconsin, Germany) type ultraviolet B (UV-B) irradiation (280–320 nm wavelength), were utilized. Two developmental stages of C. cautella; female larvae (18-day-old) and female pupae (1–2-day-old were kept, in separate 50 g plastic containers). Ten insects were placed in a cup and the cup was placed directly beneath the UV lamps at a distance of 30 cm. The female larval stages were given 0.30 g artificial diet as food to feed them. The UV was applied for 0, 2, 4, 6, and 12 h of intervals to C. cautella female larvae and female pupal stages. The 0 h treatment was assigned as a control.
2.4 Sample preparation for quantitative gene expression after UV bioassay
All of the insects were removed from the UV chamber after each exposure times. The female larvae and female pupae (whole organism) were rinsed in PBS (Phosphate Buffered Saline) (1×, pH 7.5) for 1 min before being placed in a labelled 1.5 ml Eppendorf tube and stored at −80 °C for the next step.
2.5 Total RNA extraction and vitellogenin gene expression analysis
Total RNA was extracted from UV-treated C. cautella female larvae and female pupae using Tri-RNA Reagent (Favorgen Biotech Corp., Taiwan), following the protocol strictly. The total RNA concentration was measured by using Nanodrop 2000 Spectrophotometer (Thermo Scientific, USA). After measuring the concentration, total RNA was treated with DNase to remove DNA contaminations. After that, one to two micrograms of DNase-treated RNAs from the UV-treated samples were used for cDNA synthesis. After cDNA synthesis vitellogenin gene expression was quantified using power SYBR® Green PCR Master Mix (Applied Biosystems, USA) and 7500 Fast Real-Time PCR system (Applied Biosystems, USA).
2.6 Data collection
After each treatment time, the larval and pupal mortality were counted. The female pupae emerged as adult were allowed to have mating with normal male moths in a separate plastic box and their biological characteristics were studied.
2.7 Statistical analysis
Cycle threshold of vitellogenin expression data was tabulated and means were compared between the treated and non-treated individual expression data. One-way analysis of variance was used to analyze the quantified real-time PCR data and biological studies data at (α = 0.05) (SAS, 2009).
3 Results
3.1 Larval mortality and pupal development
When the results were compared to the larval mortality in the control treatment, the UV had a detrimental effect on larvae for all exposure periods. The larval mortality was highly significant among the exposure timings (Table 1). In the control treatment, there was very little larval mortality, and the majority of the larvae developed into pupae. It was shown that mortality was significantly higher during 2 and 4 h. The difference in larval mortality between these two exposure time intervals (2 and 4 h) was not statistically significant. Mortality was directly proportional to exposure time at 6 and 12 h and became highly significant. Similar to the 2 and 4 h exposure treatments, there was no difference in larval mortality between 6 and 12 h of exposure duration. After 6 and 12 h of exposure, no larvae grow into pupae (Table 1). In each column, means followed by the same letter (s) are not significantly different, at (α = 0.05).
Parameters
Exposure (h)
Mortality (%)
ANOVA Parameters
N
F
df
P
Larval mortality
0 (Control)
0.00 ± 0.00b
3
4.50
4,14
0.0245
2
30.00 ± 30.00ab
3
4
30.50 ± 30.00ab
3
6
90.00 ± 0.00a
3
12
90.00 ± 0.00a
3
Pupal development
0 (Control)
90.00 ± 0.00a
3
4.50
4,14
0.0245
2
60.00 ± 52.00ab
3
4
60.00 ± 52.00ab
3
6
0.00 ± 0.00b
3
12
0.00 ± 0.00b
3
3.2 Ultraviolet effect on pupal mortality and adult emergence
Ultraviolet radiation had a detrimental impact on pupal survival and development at all exposure intervals, and the difference in Pupal mortality between the UV treatment and the control was extremely significant (Table 2). Some pupae died in the control treatment as well, but the large majority of pupae survived to adult. Pupal mortality was higher in the ultraviolet treatments than in the control group. There was no difference in pupal mortality after 4, 6, or 12 h of exposure (Table 2). During the UV treatment, the most of the pupae failed to mature, turned blackish in color, and died. Adults that emerged from UV-treated pupae were abnormal because their wings were not fully developed and they could not fly. Even the majority of them were unable to fully emerge from the puparium (Fig. 1). In brief, the adults were malformed and unable to continue with their biological activities. In each column, means followed by the same letter (s) are not significantly different, at (α = 0.05).
Parameters
Exposure (h)
Mortality (%)
ANOVA Parameters
N
F
df
P
Pupal mortality
0 (Control)
26.60 ± 0.00b
3
4.60
4,14
0.0229
2
47.26 ± 8.06ab
3
4
55.00 ± 4.20a
3
6
63.86 ± 13.06a
3
12
72.26 ± 8.86a
3
Adult emergence
0 (Control)
63.40 ± 0.00a
3
4.60
4,14
0.0229
2
42.73 ± 52.00ab
3
4
35.00 ± 52.00b
3
6
26.13 ± 0.00b
3
12
17.73 ± 0.00b
3

Cadra cautella adults abnormal eclosion from female pupae after various UV exposure time.
3.3 Ultraviolet effect on female moth biological characteristics
UV had a negative impact on the biological aspects of female moths that emerged from pupae. As previously stated, the majority of the UV tread pupae were deformed and unable to move. UV also has an impact on their many biological systems, including the reproductive system. Females that emerged from UV-treated pupae were allowed to mate with adult C. cautella healthy male moths. According to the results, C. cautella female moths were unable to copulate normally with male moths and could not lay enough eggs. Female moths that emerged from pupae after 2 h of UV radiation could copulate and lay eggs. Despite the fact that the female laid some eggs throughout the 4-hour treatment, none of them hatched. The overall differences between treatment and control were highly significant (Table 3). In each column, means followed by the same letter (s) are not significantly different, at (α = 0.05).
Exposure (h)*
Total laid eggs
Hatchability %
Female life span (d)
0 (Control)
184.33 ± 7.88a
60.03 ± 0.98a
7.33 ± 0.66a
2
74.33 ± 21.18bc
63.10 ± 7.05b
8.00 ± 0.00c
4
0.6 ± 0.61c
0.00 ± 0.0bc
2.66 ± 2.66bc
3.4 Ultraviolet effect on vitellogenin expression
Ultraviolet light is extremely harmful to living things. Although it may not immediately kill the animals, it has a deleterious influence on their Genetic material. In the current work, real-time PCR quantification of the vitellogenin gene confirmed that UV had an effect on the expression of this gene in C. cautella larvae, pupae, and adults. The vitellogenin expression was measured in terms of cycle threshold (CT). The CT values are inversely proportional to the target transcript detection level i.e., lower the CT value higher the amount of target transcript in the sample. In the UV-irradiated female larvae the vitellogenin expression was exhibited significantly as compared with the quantitative expression of the vitellogenin in 0 h exposure larvae. However, no increase in vitellogenin transcript was observed in female larvae exposed to UV for various durations (Fig. 2 A). The results of vitellogenin expression in pupae samples differed from the results in larvae samples, and an increase in vitellogenin expression was observed in pupae samples after 6 and 12 h of UV exposure (Fig. 2 B). However, the C. cautella female moths which emerged from the UV-treated pupae, showed the maximum down regulation of vitellogenin expression particularly the female moths which emerged from the UV-treated pupae after 6 and 12 h of exposure (Fig. 2 C).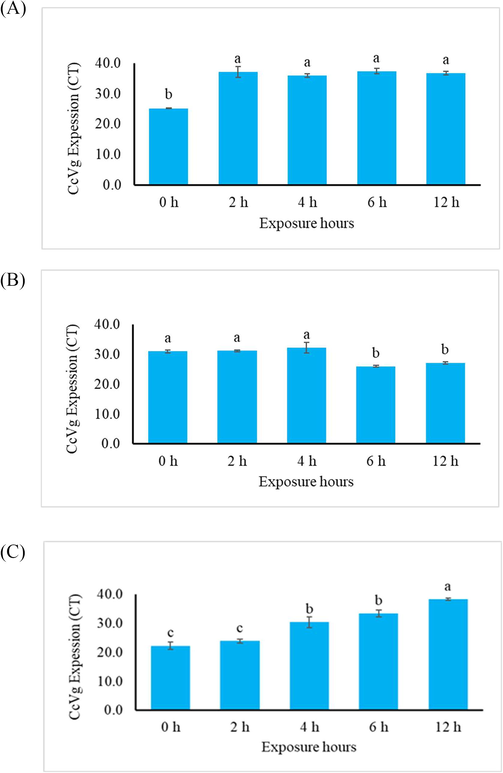
Real time PCR, cycle threshold values for the vitellogenin gene expression in Cadra cautella female larvae (1A), female Pupae (1B), and female Adult (1C), after ultraviolet exposure for different time intervals.
4 Discussion
Date fruits are vital nutritional source for human on this planet. The insect infestation threat to the products like date fruits and other grains are very challenging. The holometabolous organisms always keep growing to develop through the natural phenomenon of removing old skin and grows the new skin and enter into a new phase of development. This phenomenon is a disadvantage for them as they become very vulnerable for a specific time interval particularly, after caste off the old skin and formation of the new skin. Usually molting take place 5–6 time during the larval span. During this particular time interval, they juveniles are very delicate to be attacked for their population reduction.
Several studies have confirmed that UV can kill the larval stages of insects in the food items (Faruki et al., 2007). The C. cautella larval mortality was directly proportional to the UV exposure. During UV application it was noticed that larvae were trying to escape from the UV light and its effects which have been reported in other organisms too (Osakabe, 2021). The problem of larval escape was noticed during the experimental trial, and it may play a positive role in the storage facility during pest management utilizing UV. Because UV will be applied throughout the storage building, wherever the larvae move, they will be exposed to UV. After exposures to UV the larvae did not show any immediate symptoms of infection, however after getting out of the UV chamber they become sluggish and stop feeding at next day. The larvae that could not succeed to pupate become dark black in color and died. The UV burns the cuticle of organisms when exposed directly to it and it is one of the adverse effects of UV (Cutler and Zimmerman, 2011; Hori et al., 2014). After the UV exposure, larvae were not active and their movement was abnormal that is atypical sign of UV harmful effect on the biological functions of the living organisms. With the passage of the time of UV post exposure, the larvae become completely paralyzed and stopped moving and feeding and their body started to squeeze and turning into the blackish color.
The larval samples after each exposure preserved at −80 °C were tested for the gene expression study particularly the vitellogenin gene which is vital for the embryonic/ovarian development in oviparous organisms (Tufail et al., 2014; Li et al., 2016). UV irradiation-induced DNA damage chemically, alters DNA components which mutate the nucleotide sequence of genes (Chatterjee and Walker, 2017).
The pupae exposed to UV did not show any apparent signs of death, but their color became darker and darker with time. Under normal conditions, this shift in puparium shell color could be caused by pupal growth. It was hard to differentiate between dead and alive pupae until the adult moths emerged from the control treatment. The pupae in the UV treatment were considered dead two to three days after the adult moths emerged from the pupae in the control treatment (Alwaneen et al., 2019). Although adult eclosion occurred in certain UV-treated pupae, the adult could not have emerged fully from the pupa shell and could not fly as shown in the pictures. The post emergence physiological changes not took placed in the adults. Their wings could not get straighten after eclosion, and they were unable to copulate or show any other sign of being healthy and active.
5 Conclusion
The current study results are highly encouraging and may be useful in suppressing C. cautella populations in storages and managing this renowned pest of stored date fruits and other agricultural commodities on an industrial/commercial scale. Further detailed molecular studies might explore the effect of UV at cellular level in living organisms.
Acknowledgements
This project was funded by the National Plan for Science, Technology, and Innovation (MAARIFAH), King Abdulaziz City for Science and Technology, Kingdom of Saudi Arabia, Award Number (2-17-04-001-0044).
Disclosure of funding
The authors declare that they don’t have any particular funding for this study.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Effect of covering dates fruit bunches on Ephestia cautella Walker (Lepidoptera: Pyralidae) infestation: population dynamics studies in the field. Int. J. Agric. Appl. Sci.. 2013;5(1)
- [Google Scholar]
- Effects of temperature on the development of Ephestia cautella (Walker) (Pyralidae: Lepidoptera): a case study for its possible control under storage conditions. Pak. J. Zool.. 2013;45(6)
- [Google Scholar]
- Prediction of survival ratios of Cadra cautella (Lepidoptera: Pyralidae) different life stages after treated with ultraviolet radiation in dates. Saudi J. Biol. Sci.. 2019;26(7):1358-1363.
- [Google Scholar]
- Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen.. 2017;58(5):235-263.
- [Google Scholar]
- Ultraviolet irradiation and the mechanisms underlying its inactivation of infectious agents. Anim. Health Res. Rev.. 2011;12(1):15-23.
- [Google Scholar]
- Effects of ultraviolet (254nm) irradiation on egg hatching and adult emergence of the flour beetles, Tribolium castaneum, T. confusum and the almond moth, Cadra cautella. J. Insect Sci.. 2007;7(36):1-6.
- [Google Scholar]
- Purification and site of synthesis of Aedes aegypti yolk proteins. J. Exp. Zool.. 1972;182(3):367-377.
- [Google Scholar]
- Possible effects of ultraviolet ray (UV-C) on biological traits of Callosobruchus maculatus (Col.: Chrysomelidae) J. Stored Prod. Res.. 2016;69:91-98.
- [Google Scholar]
- Lethal effects of short-wavelength visible light on insects. Sci. Rep.. 2014;4:7383.
- [Google Scholar]
- Comparative efficacy of CO2 and ozone gases against Ephestia cautella (Lepidoptera: Pyralidae) larvae under different temperature regimes. J. Insect Sci.. 2015;15(1):126.
- [Google Scholar]
- Biological traits of Cadra cautella (Lepidoptera: Pyralidae) reared on Khodari date fruits under different temperature regimes. J. Econ. Entomol.. 2017;110(4):1923-1928.
- [Google Scholar]
- RNAi-mediated silencing of vitellogenin gene curtails oogenesis in the almond moth Cadra cautella. PloS ONE. 2021;16(2):e0245928.
- [Google Scholar]
- Influences of stored product insect movements on integrated pest management decisions. Insects. 2019;10(4):100.
- [Google Scholar]
- Molecular cloning and expression of the vitellogenin gene and its correlation with ovarian development in an invasive pest Octodonta nipae on two host plants. Bull. Entomol. Res.. 2016;106(5):642-650.
- [Google Scholar]
- Effects of modified atmosphere package (MAP) with a silicon gum film window on the quality of stored green asparagus (Asparagus officinalis L) spears. LWT - Food Sci. Technol.. 2015;60(2):1046-1053.
- [Google Scholar]
- Suppression of bedbug’s reproduction by RNA interference of vitellogenin. PloS ONE. 2016;11(4):e0153984.
- [Google Scholar]
- The use of modified and controlled atmospheres for the disinfestation of stored products. J. Pest Sci.. 2012;85(3):301-322.
- [Google Scholar]
- Vitellogenin in the eggs of the cockroach, Blattella germanica: purification and characterization. Dev. Growth Differ.. 1975;17(3):237-246.
- [Google Scholar]
- Biological impact of Ultraviolet-B radiation on spider mites and its application in integrated pest management. Appl. Entomol. Zool.. 2021;56(2):139-155.
- [Google Scholar]
- Silencing of vitellogenin gene contributes to the promise of controlling red palm weevil, Rhynchophorus ferrugineus (Olivier) Sci. Rep.. 2021;11(1):1-12.
- [Google Scholar]
- Structural characteristics of insect vitellogenins. Reprod. Biol. Invertebr.. 2002;12:69-101.
- [Google Scholar]
- SAS. 2009. SAS Institute. 2009. SAS/STAT 9.2. Users guide. SAS Institute, Cary, NC.
- Molecular characterization and expression pattern of Spodoptera litura (Lepidoptera: Noctuidae) vitellogenin, and its response to lead stress. J. Insect Physiol.. 2009;55(7):608-616.
- [Google Scholar]
- Vitellogenin of the cockroach, Leucophaea maderae: nucleotide sequence, structure and analysis of processing in the fat body and oocytes. Insect Biochem. Mol. Biol.. 2002;32(11):1469-1476.
- [Google Scholar]
- Evidence for two vitellogenin-related genes in Leucophaea maderae: the protein primary structure and its processing. Arch. Insect Biochem. Physiol.. 2007;66(4):190-203.
- [Google Scholar]
- Analysis of the vitellogenin gene of rice moth, Corcyra cephalonica Stainton. Arch. Insect Biochem. Physiol.. 2014;87(3):126-147.
- [Google Scholar]
- Simulation of heating uniformity in a heating block system modified for controlled atmosphere treatments. J. Stored Prod. Res.. 2016;65:19-29.
- [Google Scholar]
- Vitellogenin gene of the silkworm, Bombyx mori: structure and sex-dependent expression. FEBS Lett.. 1994;356(2–3):207-211.
- [Google Scholar]




