

Translate this page into:
New record of B-chromosome through meiosis in the Egyptian locust Anacridium aegyptium (Acrididae) with indication to its origin
*Corresponding author. Present address: Department of Zoology, College of Science, King Saud University, P.O. Box 2455, Riyadh 11451, Saudi Arabia Sharaf_hesham@yahoo.com (H.M. Sharaf)
-
Received: ,
Accepted: ,
This article was originally published by Elsevier and was migrated to Scientific Scholar after the change of Publisher.
Abstract
The basic chromosome complement of the Egyptian male locust, Anacridium aegyptium; collected from Giza Governorate, Egypt, is 2N = 22 + XO. All autosomes and sex chromosome are telocentric. The autosomes can be divided into three size-groups; three long (L1–L3), five medium (M4–M8) and three short chromosomes (S9–S11). The X-member is the longest member of all chromosomes. An extra telocentric B-chromosome was detected in two males of the collected locusts, nearly equal in size with S10-member and thus the chromosome complement of these males is 2N = 22 + XO + B. The origin of this extra member is discussed through its behaviour through meiosis that studied. The B-chromosome was frequently observed neighbouring the X-chromosome through most of the meiotic stages and this behaviour reinforces the present suggested hypothesis that B-chromosome was originated from X-chromosome.
Keywords
B-chromosome
Autosomes
Sex chromosomes
Meiosis

1 Introduction
Abdel-Haleem (1992) reported that the chromosome complement of male Anacridium aegyptium is composed of 23 chromosomes. He observed that autosomes and X-chromosome are more spreading through mitosis than meiosis. Thus, they observed acrocentric through mitosis stages and telocentric through meiotic ones. The chromosomes could be measured and divided into three groups, on the basis of their relative lengths, three pairs of large (L), five pairs of medium (M) and three pairs of small homologues (S). The X-chromosome is the longest member of all complement. The properties of accessory or B-chromosomes had been reviewed by Müntzing (1974). Nokkala and Nokkala (2004) declared that B-chromosomes are not belonging to the regular karyotype of any species of Acrididae.
Martin et al. (1996) found a decrease of mating involving males with B-chromosomes in the grasshopper Eyprepocnemis plorans. In the same grasshopper, Camacho et al. (1997) concluded that B-chromosome has no detectable effects on fitness. Cabrero et al. (1984) reported that the B-chromosomes in the locust Locusta migratoria are mitotically unstable, accumulated in the male germ-line and meiotically there is no significant differences in mean chiasma frequency between locusts with and without B-chromosomes. In this respect, the effect of B-chromosome on chiasma frequency in L. migratoria was studied by Viseras et al. (1988).
Bidau (1986) found a nucleolar-organising region in B-chromosome of the grasshopper Dichroplus pratensis. He also analyzed the meiotic behaviour of B-chromosome through meiosis. The same author (1988) investigated the effects of B-chromosomes on meiosis of the grasshoppers Dichroplus pseudopunctulatus and Sinipta dalmani. Henriques-Gil et al. (1989) noted accumulation of B-chromosomes in the gametes with X-chromosome in the examined males of E. plorans. Lopez-Leon et al. (1996) found that B-chromosomes were more frequent in males than females in E. plorans. Bugrov et al. (1999) stated that one B-chromosome in E. plorans has no effect on the mean chiasma frequency. They added that B-chromosomes are made up of DNA-sequences (ribosomal-DNA and a 180-bp repeat).
Abdel-Haleem (2003) studied the B-chromosome behaviour through mitotic and meiotic divisions in the grasshopper E. plorans and its relation with X-chromosome. Nokkala and Nokkala (2004) declared that B-chromosome behaviour in both mitosis and meiosis is without segregation and chiasmatic mechanism.
Therefore the main aims of the present work were to describe the effects of the supernumerary or B-chromosome in the meiotic division of the Egyptian locust A. aegyptium and to search for its origin.
2 Materials and methods
2.1 Rearing
The colony of the locust A. aegyptium was initiated by 20 males and females obtained from Abo-Roach, Giza Governorate, Cairo, in August 2007. The individuals were kept in cubic cage of 30 × 30 × 30 cm3 dimensions, at the laboratory conditions (28 ± 2 °C and 65% R.H.) and were daily fed on leaves of lettuce.
2.2 Chromosome preparation
Meiotic cells were obtained from the testes of two adult male of A. aegyptium that had B-chromosome. The testes were dissected out, cleaned and placed in insect-saline solution for a half hour, fixed in 3:1 absolute ethanol:glacial acetic acid for at least 12 h at 4 °C. Then, the fixed-testes were washed and kept in mixture of 70% ethanol (99 ml) and concentrated glycerol (1 ml) and kept at 4 °C. Then, the squash preparations were made using 4% Giemsa stain. Finally, observations were photographed using light microscopy and printed.
2.3 G-banding pattern preparation
In general, G-banded-chromosomes revealed more detailed information about chromosomes than the classical karyotype. C-banding pattern preparation was made according to the technique of Webb (1976), where the colchicine-treated-meiotic cells were immersed in trypsin solution at room temperature, washed in phosphate-buffered solution followed by immersion in 4% Giemsa stain.
3 Results
3.1 The basic complement
In A. aegyptium, the basic chromosome complement showed 11 bivalents plus X-univalent. The autosomes and X-chromosome are telocentric. The autosomes can be divided into three groups, on the basis of their relative lengths, three pairs of large (L), five pairs of medium (M) and three pairs of small homologues (S). The X-chromosome is the longest member of all chromosomes (Fig. 1(7)).
3.2 The supernumerary chromosome
A telocentric B-chromosome was found in one male of the collected locusts A. aegyptium. It is small in size and its length is slightly longer than S9-member. Thus, the chromosome complement in this male was 2N = 22 + XO + B (Fig. 1(7)). In the majority of meiotic stages, B-chromosome is neighbouring the sex chromosome (Figs. 1(1–9)). Moreover, the B-chromosome behaviour through the majority of meiotic stages revealed that it is stable element. It displays a positive heteropycnosis during leptotene, zygotene and pachytene stages. In diplotene stage, the B-chromosome observed unassociated with any chromosome. In metaphase-I, both B- and X-chromosomes are unassociated and found as univalents. In late anaphase-II, the B-chromosome was the latest-moving member towards one of the two poles, in which the sex chromosome is found, in spite of its univalent form. It seems to be approached to the pole where X-chromosome found and attached to it in an abnormal manner. In late telophase-II, it is easy to see the B-chromosome near to two out of the four haploid nuclei. It shows as small micronucleus neighbouring one of the two spermatids (Fig. 1(1)).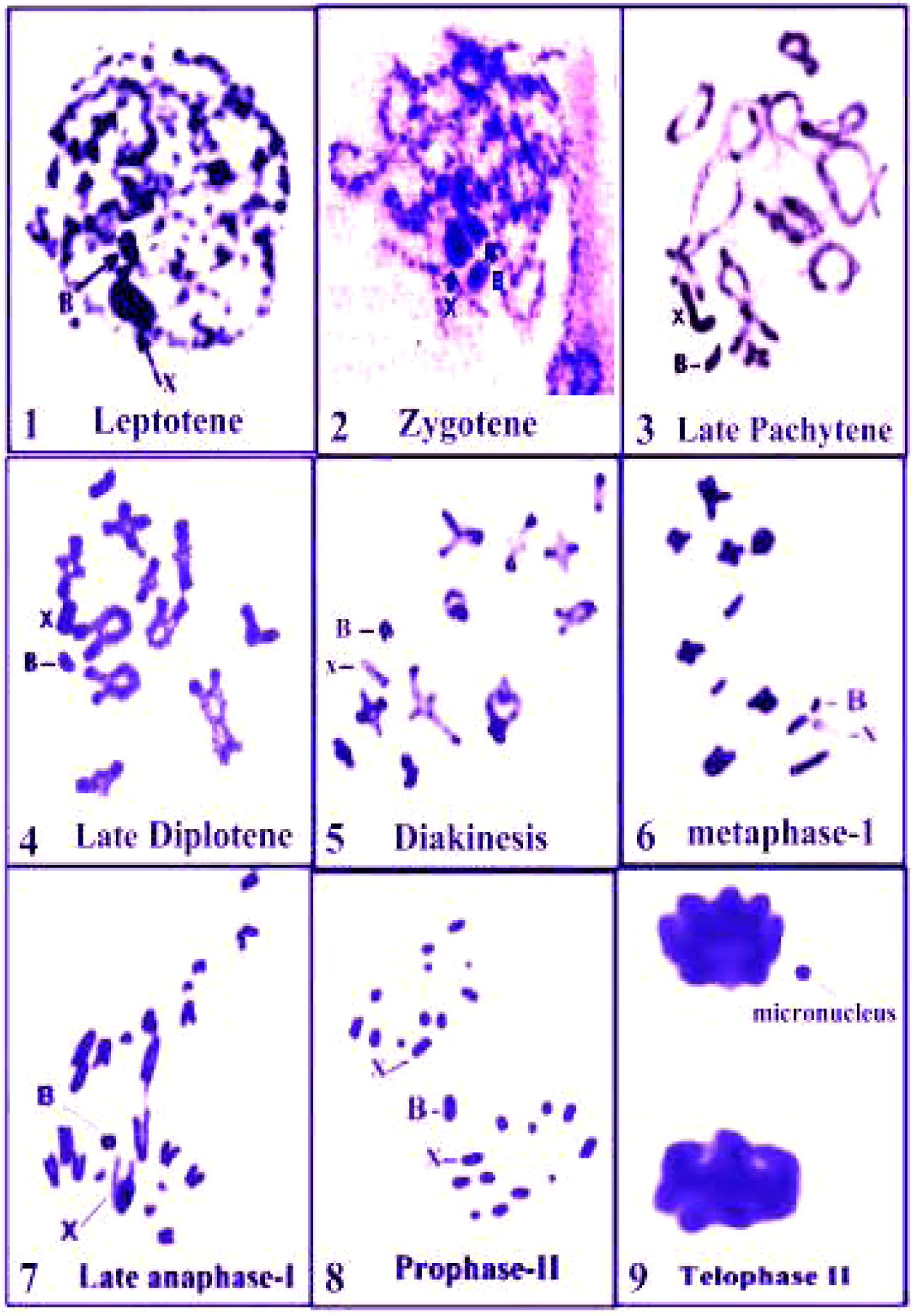
Sex chromosome behaviour of Anacridium aegyptium through meiotic prophase-1 stages, metaphase-1, anaphase-1, prophase-II and telophase-II.
4 Discussion
Bugrov et al. (1999) reported that the chromosome complement of male E. plorans is composed of 23 acrocentric chromosomes. The present result reinforced the diploid number of the same species (22 + XO).
Henriques-Gil et al. (1984) classified 14 types of B-chromosomes in the grasshopper E. plorans according to their sizes and meiotic behaviour.
In the present work, the B-chromosome of the locust A. aegyptium is telocentric slightly longer than S9-member and is a meiotically stable element. These results support those of Hewitt (1979) who declared that B-chromosomes of grasshoppers are stable in cell division. However, Bidau (1988) studied the effects of B-chromosomes in meiosis of the grasshoppers, D. pseudopunctulatus and S. dalmani and showed an irregular meiotic behaviour (lagging at the first and second meiotic divisions and finally formation of B-chromosomes as microspermatids).
In this regard, Camacho et al. (1980) displayed two B-chromosomes in E. plorans that are similar in size and both are telocentric.
Webb (1976) declared that the C-banded B-chromosome in the locust Chortoicetes terminifera was darkly stained, but such banding was not successful in identifying the origin of B-chromosome. Webb result agrees with the Present work, especially through meiotic diakinesis stage (Fig. 1(1–9)).
In the present investigation, the meiotic behaviour of the B-chromosome demonstrated that it was frequently localized near to the univalent X-chromosome and lately reached to the pole which has X-member. This behaviour supports the study of Camacho and Cabrero (1982) who found an association between the B-segment and X-chromosome in five species of grasshoppers.
On the other hand, the effect of B-chromosome on sperm formation was investigated in the present work where large numbers of B-chromosome, as micronuclei, were observed neighbouring the poles that involved X-chromosomes.
Bidau (1986) found that the B- and X-chromosomes were associated preferentially during prophase-I of the grasshopper D. pratensis and this persistent X–B-association leads to both chromosome moving to the same pole through anaphase-I. The present results in A. aegyptium support that persistent X–B-association.
Bugrov et al. (1999) showed that the B-chromosomes of male E. plorans are made up of the same two DNA-sequences of X-member (ribosomal-DNA and a 180-bp repeat). This result offers a strong support for the present hypothesis about the origin of B-chromosome in A. aegyptium.
In this respect, Bugrov et al. (2003) analyzed the distribution of repetitive DNA of B-chromosomes of Podisma sapporensis (Acrididae), using meiotic chromosome microdissection followed by DOP-PCR (degenerated oligo-nucleotide primed polymerase chain reaction) and FISH analysis, and found the same two DNA-sequences of Bugrov et al. (1999).
Abdel-Haleem (2003) analyzed the behaviour of B-chromosome of male grasshopper E. plorans and suggested the same hypothesis that B-segment was originated from X-chromosome through mitosis and meiosis.
Furthermore, Nokkala and Nokkala (2004) analyzed the behaviour of B-chromosomes through mitosis and meiosis and found that they were without segregation and chiasmatic mechanism.
Warchałowska-Sliwa et al. (2005) studied the orthopteran Tetrix tenuicornis, which collected from polluted area with heavy metal in Poland, and found cytogenetic abnormalities involving anomalies in chromosome number and morphology through mitosis and meiosis, frequent presence of B-chromosome and a heterozygous deficiency in L2 and M3 bivalents in male as well as tetraploid, and octaploid chromosome numbers in the females.
References
- Abdel-Haleem, A.A., 1992. Comparative cytogenetical studies on the locusts Schistocerca gregaria and Anacridium aegyptium. M.Sc. Thesis, Faculty of Science, Ain Shams University, Cairo, pp. 33–34.
- B-chromosome behaviour through mitotic and meiotic divisions of male Egyptian grasshopper Eyprepocnemis plorans plorans and its relation with sex chromosome. J. Egypt. Ger. Soc. Zool.. 2003;42:147-156.
- [Google Scholar]
- A nucleolar-organizing B-chromosome showing segregation-distortion in the grasshopper Dichroplus pratensis (Melanoplinae: Acrididae) Can. J. Genet. Cytol.. 1986;28(1):138-148.
- [Google Scholar]
- Microspermatid and macrospermatid production induced by meiotically unstable B-chromosomes in two species of grasshoppers (Acrididae) Rev. Bras. Genet.. 1988;11(3):547-562.
- [Google Scholar]
- Karyotypic features of Eyprepocnemidinae grasshoppers from Russia and Central Asia with reference to the B-chromosomes in Eyprepocnemis plorans (Charp) Folia Biol.. 1999;47(3):97-104.
- [Google Scholar]
- B-chromosomes of the Podisma sapporensis (Orthoptera, Acrididae) analysed by chromosome microdissection and fish. Folia Biol.. 2003;51(12):1-11.
- [Google Scholar]
- The B-chromosomes of the locust Locusta migratoria. I – Detection of negative correlation between mean chiasma frequency of the rate of accumulation of the B a reanalysis of the available data about the transmission of these B-chromosomes. Genetica (The Hague). 1984;64(3):155-164.
- [Google Scholar]
- Supernumerary segment in 5 species of grasshoppers (Orthoptera, Acridoidea) Genetica (Hague). 1982;59(2):113-118.
- [Google Scholar]
- B-chromosome system of the grasshopper Eyprepocnemis plorans. Chromosoma (Berlin). 1980;80(2):163-176.
- [Google Scholar]
- Population dynamics of a selfish B-chromosome neutralized by the standard genome in the grasshopper Eyprepocnemis plorans. Am. Nat.. 1997;149(6):1030-1050.
- [Google Scholar]
- Evolution of a complex B-chromosome polymorphism in the grasshopper Eyprepocnemis plorans. Chromosoma (Berlin). 1984;89(4):290-293.
- [Google Scholar]
- Sex-dependent meiotic behavior of B-chromosomes in the grasshopper Eyprepocnemis plorans. Heredity. 1989;62:217-221.
- [Google Scholar]
- Hewitt, G.M., 1979. Grasshoppers and Crickets. Animal Cytogenetics, Insecta I. Orthoptera, vol. 3. Berlin, Stuttgart, Gebri der Borntraeger.
- Achiasmate segregation of X- and B-univalents in males of the grasshopper Eyprepocnemis plorans is independent of previous association. Chromosome Res.. 1996;4(1):43-48.
- [Google Scholar]
- The effects of B-chromosomes on mating success of the grasshopper Eyprepocnemis plorans. Genetica. 1996;97(2):197-203.
- [Google Scholar]
- Interaction of B-chromosomes with A- or B-chromosomes in segregation in insects. Cytogenet. Genome. Res.. 2004;106(24):394-397.
- [Google Scholar]
- The B-chromosome of Locusta migratoria. II – Effects on chiasma frequency. Genome. 1988;30(2):118-123.
- [Google Scholar]
- Heavy metal accumulation heat shock protein expression cytogenetic changes in Tetrix tenuicornis (L.) (Tetrigidae, Orthoptera) from polluted areas. Environ. Pollut.. 2005;133(2):373-381.
- [Google Scholar]
- Chromosome organisation in the Australian plague locust, Chortoicetes terminifera. Chromosoma. 1976;55(3):229-246.
- [Google Scholar]




